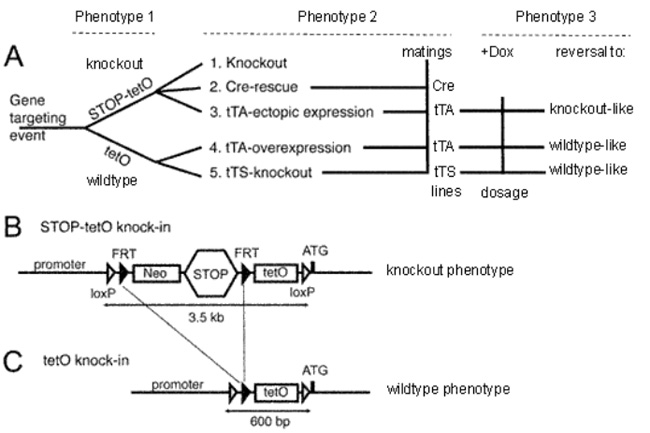
図1. F.A.S.T.™ システムの概要
F.A.S.T.™ システムでは STOP-tetO ノックインマウスとtetO ノックインマウスから 5 種類の遺伝子操作モデルを作製可能です (Fig. A)。
まずES細胞の目的遺伝子の翻訳開始部位のすぐ上流に 「loxP-FRT-Neo-STOP-FRT-TetO-loxP」 カセットを挿入することで、ノックアウト表現型を得ます (STOP-tetO ノックインマウス、Fig B)。
このマウスにフリッパーゼ (FLP) 発現または一過性発現マウスを交配することで STOP が除かれ、野生型と同様の発現パターンを示す tetO ノックインマウスを得ることができます (Fig C)。 この2つのノックインマウス (STOP-tetOとtetOマウス) に、さらにCreリコンビナーゼマウス、tTAマウス、または tTSマウスを交配することで、5種類の遺伝子操作モデルを得ることができます (Fig. A, Phenotype 2)。またドキシサイクリン (Dox) の投与により、遺伝子発現を可逆的に制御することが可能です (Fig. A, Phenotype 3)。
用語説明
DNA 組換え酵素 Cre が2つの loxP と呼ばれる DNA 配列間で部位特異的組換え反応を起こすことを利用した実験系。loxP同士の向きが逆であれば挟まれた配列は反転するが、同じ向きだと挟まれた配列が切り出される。目的遺伝子の前後にloxPを挿入したFloxed マウスと組織特異的プロモーターの制御下でCreを発現するCre マウスを交配することで、組織特異的又は時期特異的な遺伝子不活化が可能。
出芽酵母由来の組換え酵素 FLP は FRT 配列を認識して組換えを起こすため、 Cre-loxP システムと同様の目的で頻用され、主に抗生物質耐性マーカーの除去に使用されてきました。
Tetシステムとは抗生物質テトラサイクリン誘導体であるドキシサイクリン (Dox) を投与することで可逆的に目的遺伝子の発現を調節できる実験系です。このシステムは大腸菌テトラサイクリン耐性オペロンで働くTetリプレッサー(TetR)とTetオペレーター配列(tetO配列)を利用し、TetR は Dox 非存在下で tetO 配列に結合するが、Dox が結合すると tetO 配列に結合できなくなるという性質を利用しています。
テトラサイクリン調節性トランス活性化因子 (tTA) は TetR とヘルペスウイルス由来のVP16 転写活性ドメイン (VP16AD) との融合タンパク質でありtetO 配列に結合すると下流のプロモーターを活性化します。Doxと結合するとTetR が tetO 配列に結合できなくなるため tetO 配列下流のプロモーターは活性化しません。
テトラサイクリン調節性の転写サイレンサー(tTS) は、TetR と KRAB サイレンシングドメイン (SDKid-1) を融合したもので、Dox 非存在下でtetO配列に結合し、遺伝子発現を抑制します。
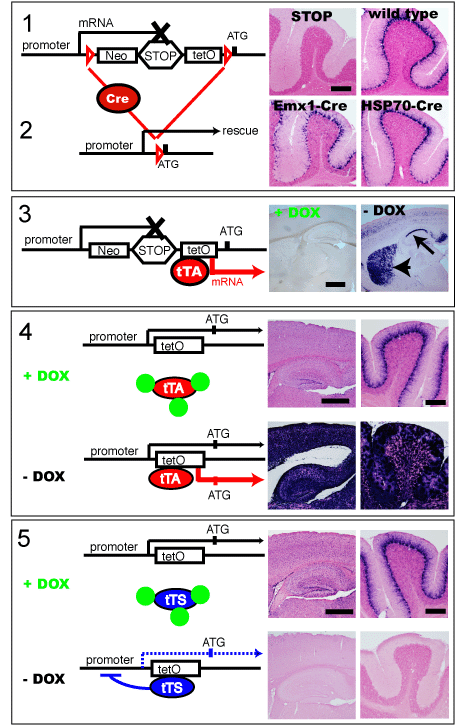
図2. Mlc1 遺伝子での 5 つの F.A.S.T.™ モデル作製例
- Mlc1遺伝子はグリア細胞に発現する遺伝子で、青紫色に染色された部分でMlc1遺伝子が発現しています。STOP-tetOノックインマウスは、野生型マウスと比較してMlc1遺伝子の発現がノックアウトされた表現型となります。
- STOP-tetO マウスと Cre マウスとの交配によりF.A.S.T.™カセットを除くことで、ノックアウト表現型はレスキューされます。
- STOP-tetOマウスと alphaCamKII-tTA マウス (神経細胞でのみ tTA を発現) を交配すると、Dox投与によりグリア細胞のシグナルが見られなくなりますが、Dox 非存在下では神経細胞 (矢印部分) でのみ Mlc1 を発現する異所性発現モデルとなります。
- tetOノックインマウスはグリア細胞でのみ tTA を発現する tTA マウスとの交配により、Dox 存在下では野生型の発現となり (上段)、Dox 非存在下では tTA が tetO 配列に結合することで Mlc1 を過剰発現することができます (下段)。
- tetOノックインマウスと tTS マウスの交配により、Dox の存在/非存在下で Mlc1 の条件的野生型発現 (上段) 及びノックアウトモデル (下段) を作製できます。























 このページを印刷する
このページを印刷する










 中身を見る
中身を見る 中身を見る
中身を見る 中身を見る
中身を見る
 中身を見る
中身を見る