






















 このページを印刷する
このページを印刷するAgrisera社では、さまざまな植物にて交差性を確認済みの細胞膜H+ATPase抗体を提供しています。植物細胞膜マーカー抗体としておすすめです。
記事ID : 45275
植物の細胞膜プロトンポンプ(H+ATPase)に対する抗体 H+ATPase抗体 (植物細胞膜マーカー抗体)
背景
細胞膜H+ATPase (H+-ATPase) は、植物や菌類の細胞膜にのみ存在すると考えられている 100 kDa のタンパク質ファミリーです。このタンパク質は生体膜内に固定されており、エネルギー源として使用される電気化学勾配を生成し、ほとんどの代謝産物の取り込みや、葉の動きなどの環境に対する植物の反応に不可欠です。
特長
| 品番 | AS07 260 |
|---|---|
| 免疫動物 | Rabbit |
| Clonality | Polyclonal |
| 免疫原 | Arabidopsis thaliana を含む、入手可能な双子葉および単子葉植物、シダ、コケ、および藻類の原形質膜 ATPase 配列に由来。 ATPase 1 (UniProt: P20649, TAIR: At2g18960) and ATPase 2 (UniProt: P19456, TAIR: At4g30190), 3 (UniProt: P20431, TAIR: At5g57350), 4 (UniProt: Q9SU58, TAIR: At3g47950), 6 (UniProt: Q9SH76, TAIR: At2g07560), 7 (UniProt: Q9LY32, TAIR: At3g60330), 8 (UniProt: Q9M2A0, TAIR: At3g42640), 9 (UniProt: Q42556, TAIR: At1g80660), 11 (UniProt: Q9LV11, TAIR: At5g62670) of Arabidopsis thaliana and hydrogen ATPase of Chlamydomonas reinhardtii (Q9FNS3) |
| 適用 | Immunofluorescence (IF), Immunolocalization (IL), Western blot (WB) 1 : 600-1 : 1000 (IF), 1 : 100 (IL), 1 : 1000-1 : 10 000 (WB)* |
| 分子量 (予想 |見かけ) |
90-95 kDa (Arabidopsis thaliana, depending upon an isoform) |
| 交差性 (確認済み) |
Actinidia chinensis, Aesculus hippocastanum, Arabidopsis thaliana, Camellia sinensis cv. Shu-chazao, Chara australis R.Br, Chlamydmonas reinhardtii, Cucumis sativus, Cucurbita moschata, Glycine max, Kandelia obovata, Hordeum vulgare, Lolium perenne, Lycopersicon esculentum, Malus x domestica Borkh. c.v. Fuji, Marchantia polymorpha, Medicago truncatula, Nicotiana benthamiana, Nicotiana tabacum, Noccaea caerulescens, Oryza sativa, Petunia hybrida, Phalenopsis Sogo Yukidian cultivar V3, Physcomitrium patens, Picea abies, Pisum sativum, Populus tremula, Pteris vittata (fern), Ricinus communis, Spinacia oleracea, Solanum lycopersicum, Tagetes erecta, Tetraselmis chuii, Zea mays, Vicia faba |
| 交差性 (予想) |
Algae, Amaranthus hypochondriacus,Avena sativa,Beta vulgaris, Cyanidioschyzon merolae, Dunaliella spp., Gossypium hirsutum, Hordeum vulgare, Ostreococcus spp., Pinus thunbergii, Physocomitrella patens, Mesembruanthemum crystallinum, Mortierella elongata, Nannochloropsis gaditana CCMP526, Ostreococcus tauri, Prosopis alba, Saccharomyces cerevisiae, Solanum tuberosum, Sorghum bicolor, Spinacia oleracea, Triticum aestivum, Ulva prolifera, Ustilago maydis |
| 非交差性 | Allium sp., Aspergillus niger, Citrus limon, Colobanthus apetala, Cuminium cyminum, Curcuma amada, Deschampsia antractica, Lupinus luteus, Morinda citrifolia, Trigonella foenum, Vicia faba |
【留意事項】
*サンプルを 70°C 以上に加熱しないでください。これにより、H+ATPase が沈殿し、ウェスタンブロットでシグナルが得られなくなる可能性があります。
SDS-PAGE の前に、サンプルを室温、10,000 rpm/1 分間で遠心分離し、凝集物を除去します。
H+ATPase は成熟した根や葉にはあまり存在しないため、検出には非常に高感度な試薬の使用が必要になる場合があります。
商品データ
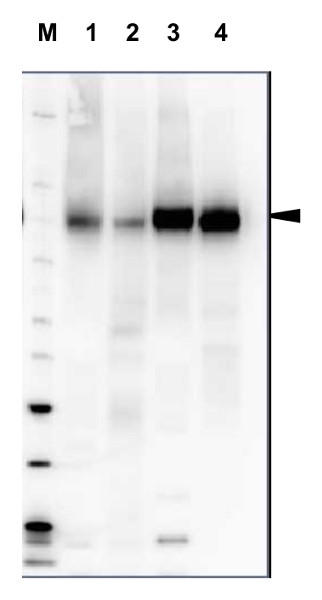
図1. さまざまな植物におけるH+ATPase検出(WB)
Lane(1) Arabidopsis thaliana, 2.0 µg (2)Hordeum vulgare, 2.0 µg, (3)Zea mays, 2.0 µg, (4)Nicotiana tabaccum plasma membrane fraction, 2.5 µg
各植物サンプルから PEB (protein extraction buffer, 品番:AS08 300) で抽出し、70℃で10分間変性後、NuPage (Invitrogen社) LDS-PAGEにアプライし、PVDF膜に転写した。ブロッキングの後、1次抗体、2次抗体と反応させ、化学発光検出試薬を用いて、シグナルを検出した。
1次抗体:抗H+ATPase抗体 (品番:AS07 260、1:5000、1 時間)
2次抗体:抗ウサギ二次抗体HRP標識 (推奨二次抗体 品番:AS09 602、1:20,000、1 時間)
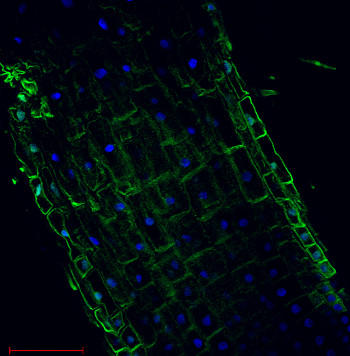
図2.Arabidopsis thaliana (根)における細胞膜 H+ATPase 局在
Arabidopsis thaliana 伸長帯中のH+ATPase (緑色)。
DAPI との共染色により核 (青色) を可視化。
Arabidopsis thaliana の根をパラホルムアルデヒドで 30 分間固定し、免疫局在化の前に組織洗浄が行われている。
1次抗体:抗H+ATPase抗体 (品番:AS07 260、1:300)
2次抗体:抗ウサギIgG抗体Alexa 555
スケールバー - 100 µm
Taras Pasternak博士, (Freiburg University, Germany)のご厚意により提供
H+ATPase抗体
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
Anti H+ATPase, Arabidopsis thaliana (Rabbit)  |
AGR | AS07 260 | 50 UG |
¥107,000 |
| Anti H+ATPase, Arabidopsis thaliana (Rabbit) |
AGR | AS07 260 | 2*50 UG |
販売終了 |
| Anti H+ATPase, Arabidopsis thaliana (Rabbit) |
AGR | AS07 260 | 3*50 UG |
販売終了 |
関連商品
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
| 1 | PEB (4x) | protein extraction buffer |
AGR | AS08 300 | 5*2 ML |
¥40,000 |
| Anti IgG (H&L), Rabbit (Goat) Horseradish Peroxidase |
AGR | AS09 602 | 1 MG |
¥71,000 |
| Anti IgG (H&L) - trial sample, Rabbit (Goat) Horseradish Peroxidase |
AGR | AS09 602-TRIAL | 10 UL |
¥8,000 |
製品使用文献(一部)
- Michalopoulou et al. (2022) The host exocyst complex is targeted by a conserved bacterial type-III effector that promotes virulence. Plant Cell. 2022 May 30:koac162. doi: 10.1093/plcell/koac162. Epub ahead of print. PMID: 35640532.
- Hofmann, Wienkoop & Luthje (2022) Hypoxia-Induced Aquaporins and Regulation of Redox Homeostasis by a Trans-Plasma Membrane Electron Transport System in Maize Roots. Antioxidants (Basel). 2022 Apr 25;11(5):836. doi: 10.3390/antiox11050836. PMID: 35624700; PMCID: PMC9137787.
- Huang et al (2021). Parasitic modulation of host development by ubiquitin-independent protein degradation. Cell. 2021 Sep 30;184(20):5201-5214.e12. doi: 10.1016/j.cell.2021.08.029. Epub 2021 Sep 17. PMID: 34536345; PMCID: PMC8525514.
- Lapshin et al. (2021) Sterol Extraction from Isolated Plant Plasma Membrane Vesicles Affects H+-ATPase Activity and H+-Transport. Biomolecules. 2021 Dec 16;11(12):1891. doi: 10.3390/biom11121891. PMID: 34944535; PMCID: PMC8699270.
- Choi et al. (2021) Augmented CO2 tolerance by expressing a single H+-pump enables microalgal valorization of industrial flue gas. Nat Commun. 2021 Oct 18;12(1):6049. doi: 10.1038/s41467-021-26325-5. PMID: 34663809; PMCID: PMC8523702.
商品は「研究用試薬」です。人や動物の医療用・臨床診断用・食品用としては使用しないように、十分ご注意ください。
※ 表示価格について














