このページを印刷する
このページを印刷するCRISPR-Cas システムには、クラス 1 とクラス 2 の 2 つのクラスが知られています。クラス 1 システムは複数のサブユニットからなるエフェクター複合体の存在を特徴とし、クラス 2 の機構は単一タンパク質のエフェクターとともに作用します。SignalChem Diagnostics社 では、クラス 2 に属する組換え Cas9、12、13 と、クラス 1 の CRISPR type III-A システムのエフェクター複合体リボヌクレアーゼとは別の酵素 Csm6 を提供しています。
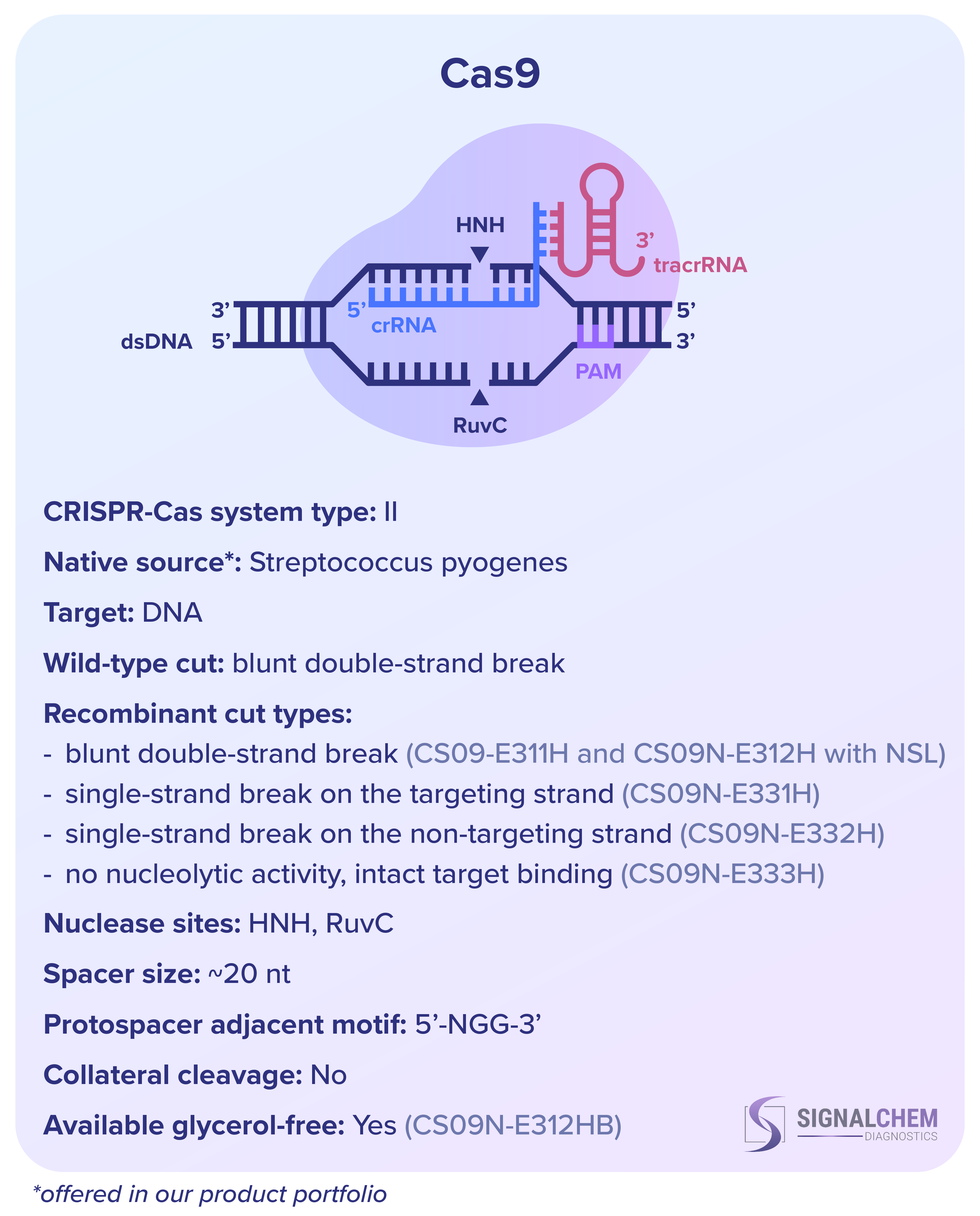
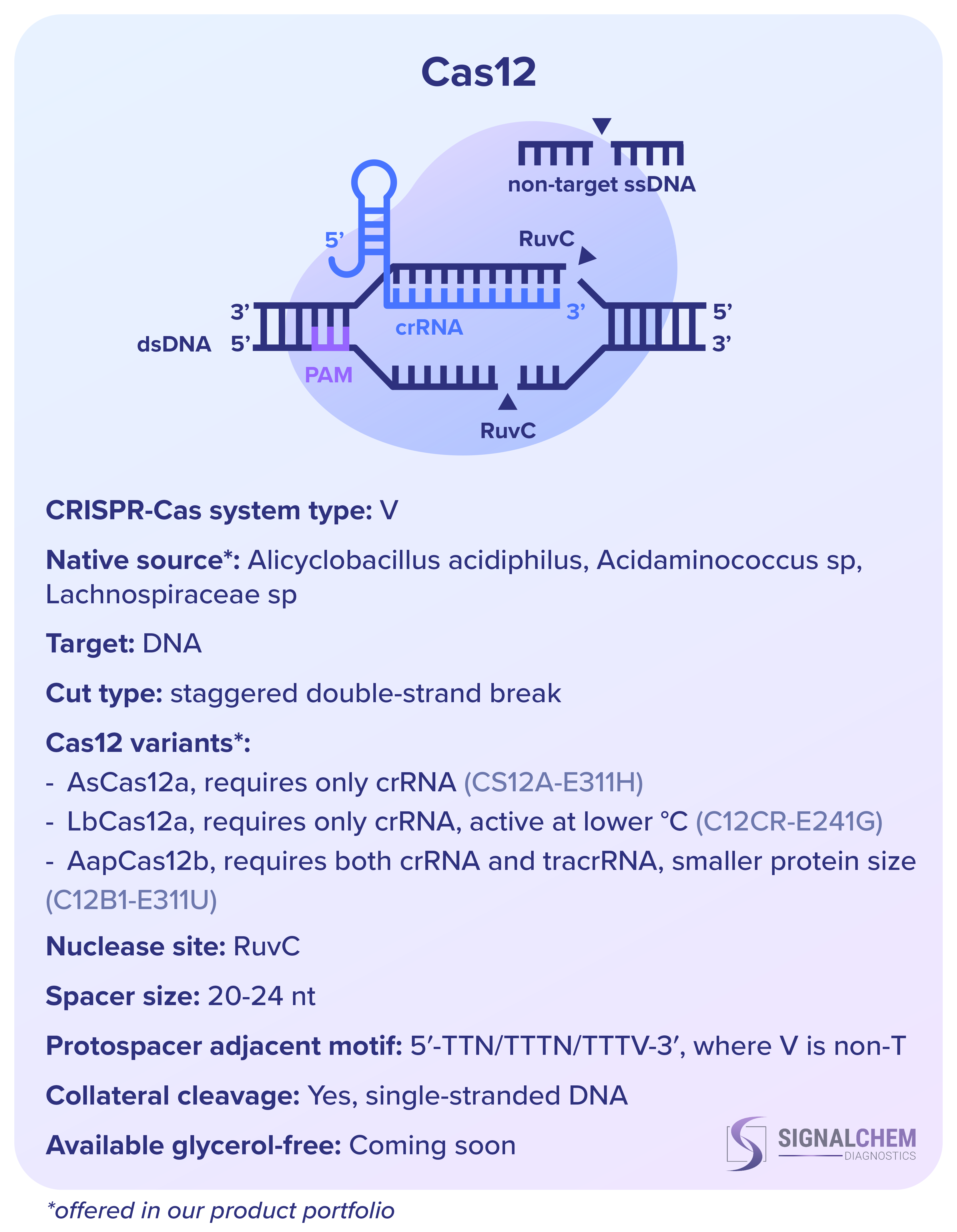
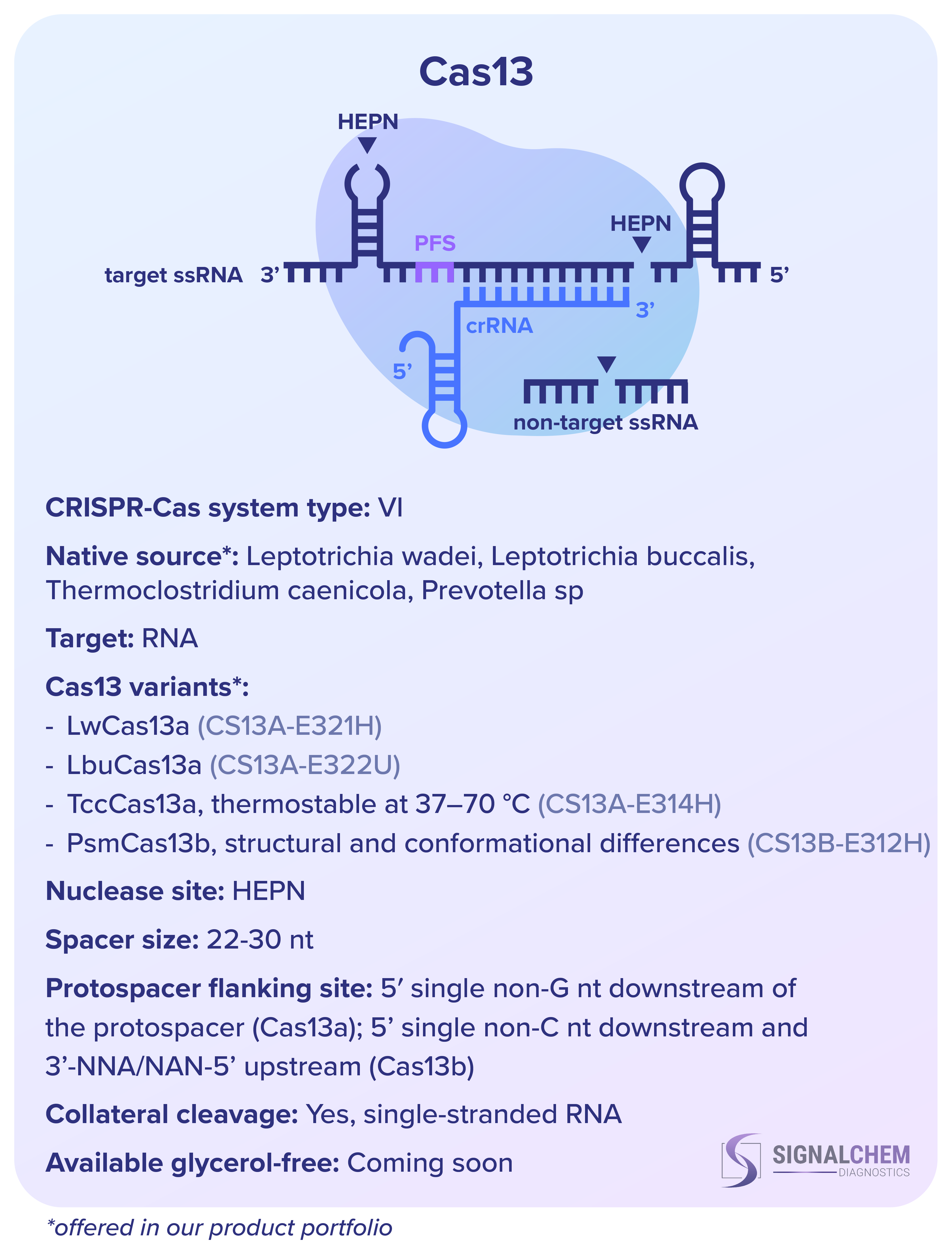
SignalChem Diagnostics社 の組換え Cas タンパク質のうち、Cas9 と Cas12 は両方とも DNA を標的としますが、RNA 成分と切断の方法、ヌクレアーゼドメイン、プロトスペーサー隣接モチーフ (PAM) の認識部位が異なります。プロトスペーサーは標的の DNA または RNA 配列であり、スペーサーは CRISPR RNA (crRNA) の一部を構成するそれに相補的な配列です。PAM とスペーサーはターゲットの認識に不可欠です。
野生型Cas9には、crRNAに加えて、その結合足場として機能するトランス活性化RNA(tracrRNA)を必要とします。しかし、in vitroでは、crRNA と tracrRNA は 1 つの単一ガイド RNA (sgRNA) に結合されることがよくあります。しかし、Cas12 は tracrRNA を必要とせず、二本鎖 DNA と一本鎖 DNA の両方を特異的に標的にすることができます。さらに、Cas12 は活性化されると、周囲の非特異的な一本鎖 DNA も切断します。
Cas9 とは対照的に、Cas13 は RNA を標的とし、tracrRNA を必要としません。補完的なものによって活性化されると、Cas13 は副次的な切断特性を示し、スペーサー配列と一致するかどうかに関係なく、周囲の RNA 分子を無差別に切断します。
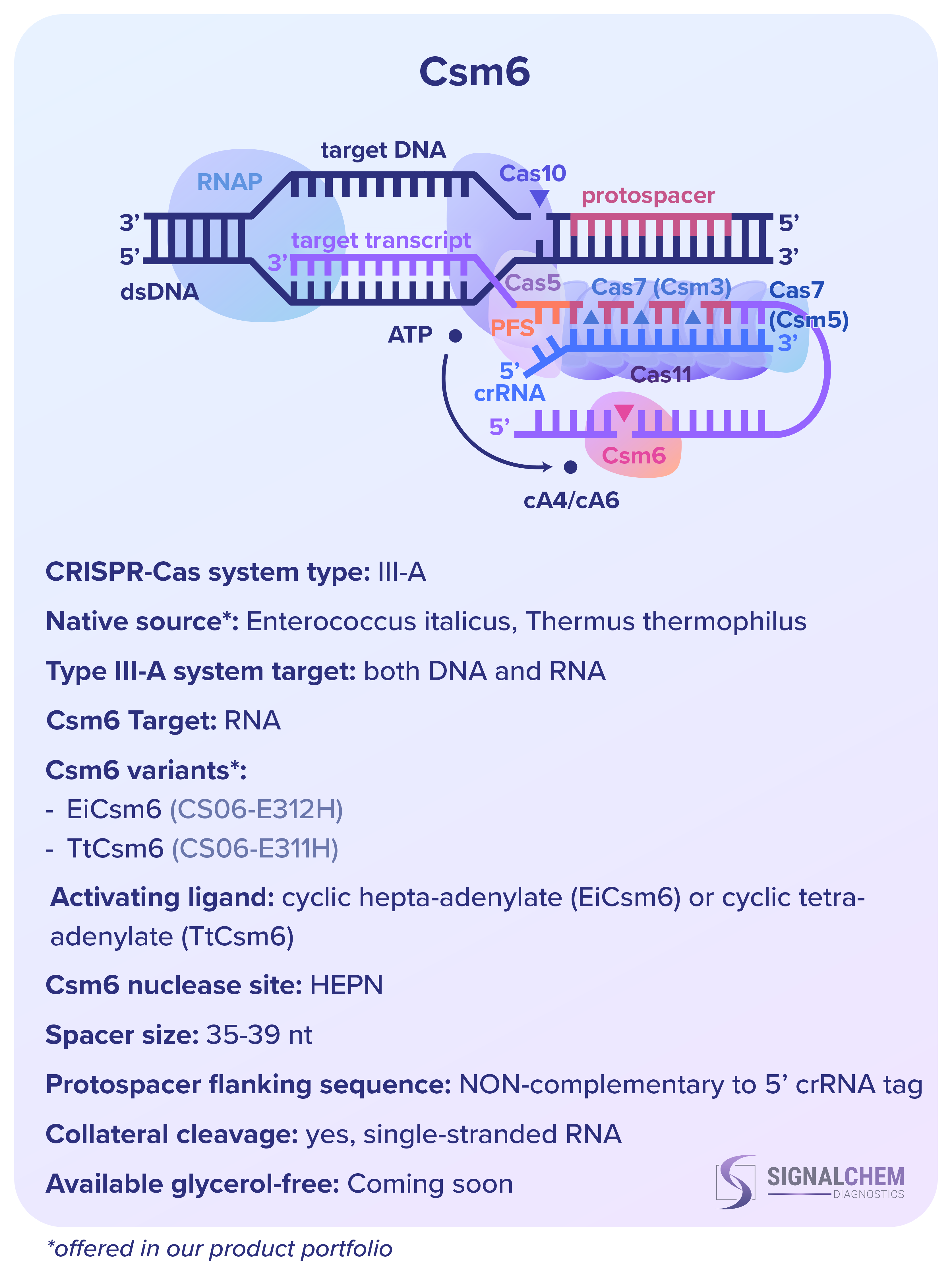
クラス 2 システムの単一タンパク質エフェクターが比較的単純であるのに対して、クラス 1 の CRISPR-Cas type III-A 機構は非常に複雑です。つまり、DNA と RNA の両方をターゲットにしており、そのエフェクター複合体はいくつかの Cas タンパク質で構成されています。
- Cas5 および Cas7 (Csm3 および Csm5 サブユニット) は crRNA をを取り込み、標的転写物への結合を促進します。その後、Csm3 サブユニットが 6 番目のヌクレオチドごとにそれを切断します。
- Cas11 の小さなサブユニットは、エフェクター複合体の頭部と尾部を接続しています。
- Cas10 サブユニットは、複合体が標的 RNA に結合して活性化された後、標的 DNA を切断します。
Cas10 活性の副産物として、Csm6 の活性化リガンドとして機能する環状オリゴアデニレートが生成されます。次に、Csm6 は一本鎖 RNA を非特異的に切断します。
CRISPR タイプ3-A システムでは、crRNA の 5' タグが転写産物のプロトスペーサー隣接配列に高度に相補的である場合、Cas10 の不活性化によって自己標的化が回避され、この領域で広範な塩基対が形成されます。ただし、このメカニズムはスペーサーが RNA プロトスペーサーと一致したとしても、標的 RNA の切断を不活性化することはありません。