このページを印刷する
このページを印刷する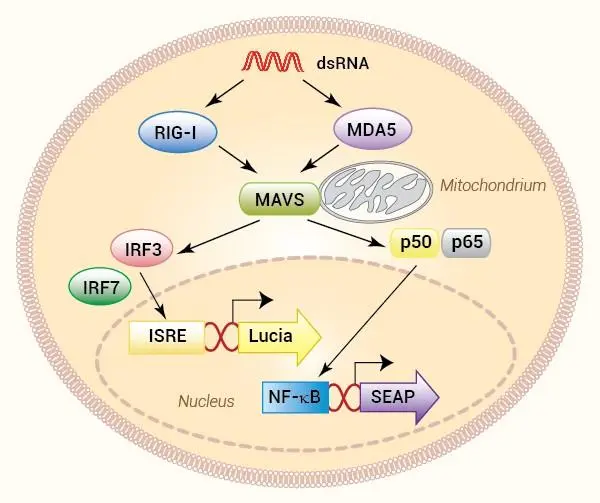
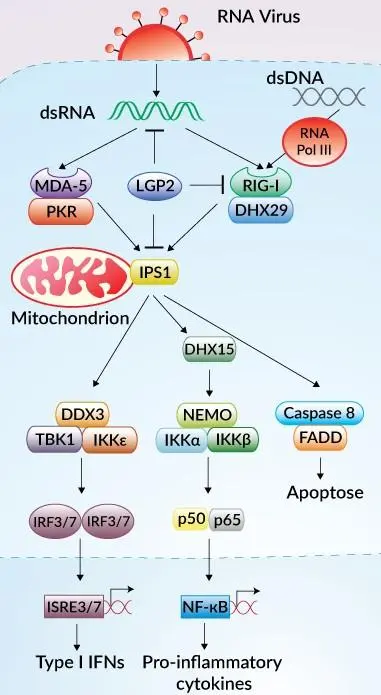
RLR(RIG-I様受容体:主に RIG-I、MDA5)は、細胞質 RNA を感知することでウイルスに対する効果的な免疫応答に不可欠です。
RIG-I または MDA5 が活性化されると、最終的に IRF3 および IRF7 依存性の I 型インターフェロン産生と、NF-κB 依存性の炎症性サイトカイン産生を引き起こします。
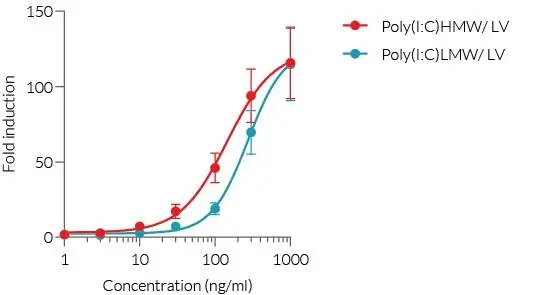
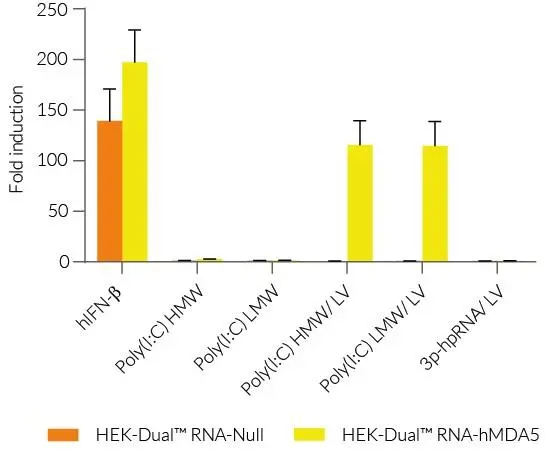
InvivoGen は、RLR(RIG-I様受容体)を介したシグナル伝達経路を容易にモニタリングするための、さまざまな ヒトおよびマウス細胞株 を提供しています。
本レポーター細胞は、NF-κB や IRF 等の転写因子により誘導されるレポーター遺伝子を発現しており、活性化に応答して SEAP(分泌型アルカリホスファターゼ) および/または Lucia(分泌型ルシフェラーゼ) を発現するように設計されています。両方のレポータータンパク質は培養上清に分泌されるため、検出試薬を用いてシグナル経路の活性化を容易に検出できます。
RLR(RIG-I様受容体)レポーター細胞株 商品一覧 は<目次>の 商品選択ガイド からご確認ください。