






















 このページを印刷する
このページを印刷する1996年、ショウジョウバエのTollタンパク質が、微生物感染症に対する防御に重要であることが発見されました1)。その1年後、現在はトールライクレセプター4(TLR4)として知られているヒトのホモログがデータベース検索により同定されました2)。現在までに、マウスで13種類、ヒトで10種類のTLRが同定されています。TLRによって誘導されるシグナル伝達は、インターフェロン(IFNs)や腫瘍壊死因子(TNF)-α、インターロイキン(IL)-6、IL-12などの様々なサイトカインの生産させる細胞応答を引き起こします。TLRの細胞質領域は、Toll/IL-1レセプター(TIR)ドメインで構成され、IL-1レセプターファミリーと高い相同性を示します。細胞外領域は、主にロイシンリッチリピート(LRRs)から成り、PAMPsの認識に重要な役割を果たしています。
TLRの発現は、樹状細胞(DCs)、マクロファージ、B細胞のような抗原提示細胞だけでなく、特定のT細胞と非免疫性細胞(例えば線維芽細胞と上皮細胞)でも見られます。これらは、バクテリア、ウイルス、菌類、原生動物由来のPAMPsを検出します。各種TLRsは、それらが認識するPAMPsによって大まかに分類されています。TLR1、TLR2、TLR6はリポペプチドを検出し、TLR3、TLR7、TLR8、TLR9は核酸を認識します。TLR5はフラジェリンを認識し、TLR4は様々なリポ多糖類(LPS)を認識します。細胞外領域とLRRsは、直接的又は間接的にリガンドと相互作用します。例えば、MD-2、CD14とLPS結合タンパク質(LBP)はTLR4のコレセプターである一方、フラジェリンはTLR5と、脱メチル化CpGオリゴヌクレオチドはTLR9と直接相互作用すると考えられています3-5)。
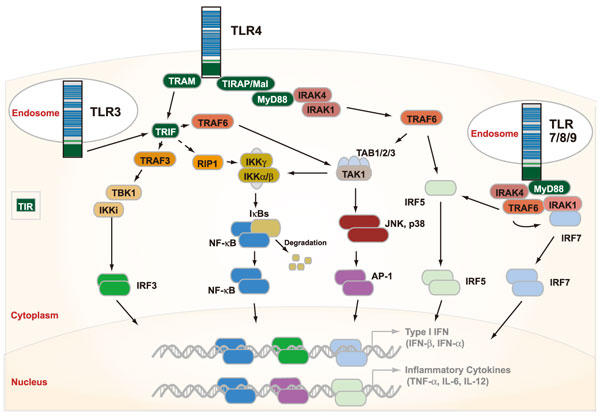
図1.TLRのシグナル経路
TLRリガンドの結合により、細胞質TIRドメインが近接してTLRがヘテロ又はホモ二量体化し、補助因子を召集します。TLRの補助因子はTIRドメインを含む分子で、MyD88、Mal(MyD88アダプター様タンパク質、TIRAP;TIR-関連タンパク質としても知られる)、TRIF(TIRドメイン含有アダプタータンパク質-IFN-β誘導、TICAM-1;TIR含有アダプター分子-1)、TRAM(TRIF関連アダプター分子、TICAM-2;TIR含有アダプター分子-2)、SARMs(sterile alpha and HEAT-armadillo motifs)などがあります。個々のTLRsにより異なるシグナル応答が誘導されることの一因として、このような様々な補助因子を利用していることが考えられます。
Cosmo Bio would like to acknowledge and thank the Enzo Life Sciences,Inc. for providing TLR information presented here.














