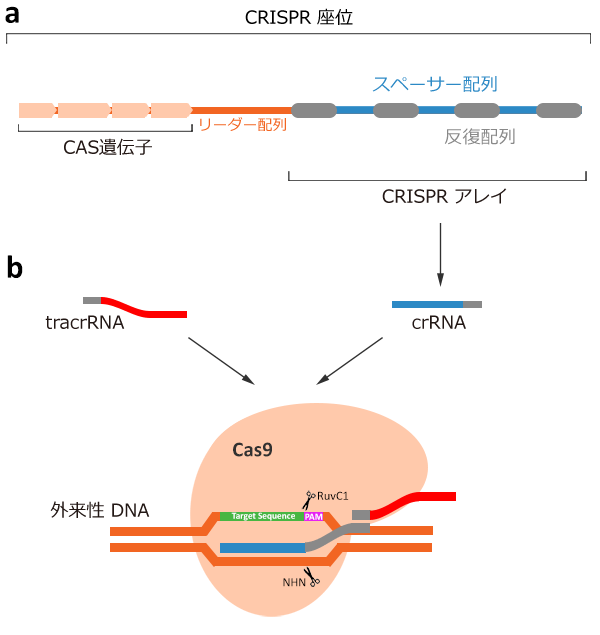
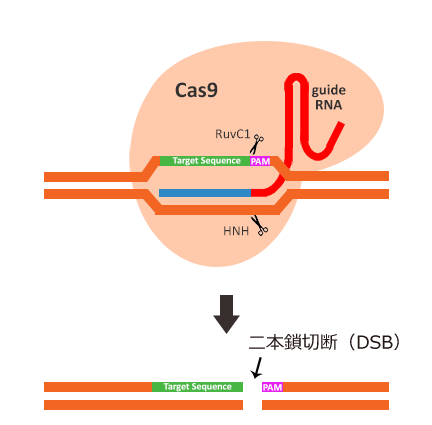
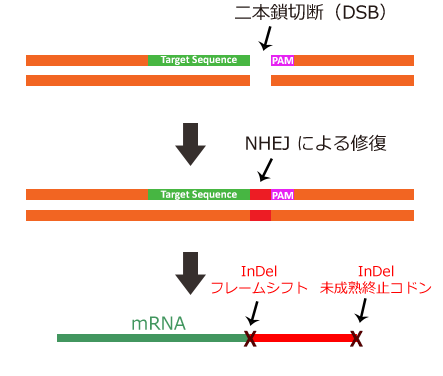
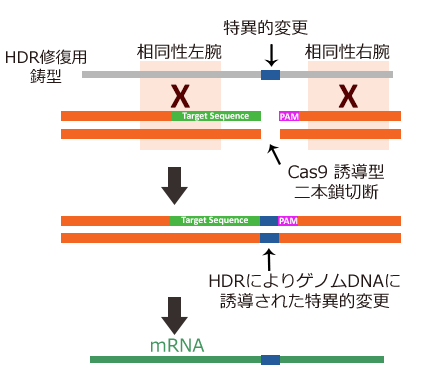
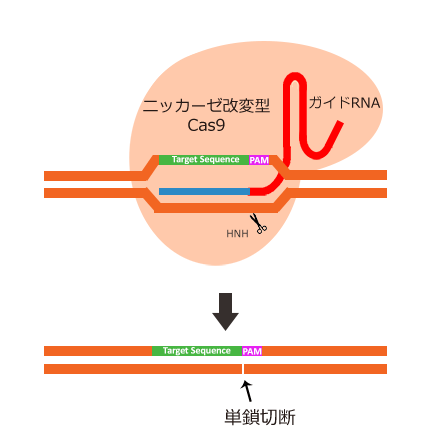
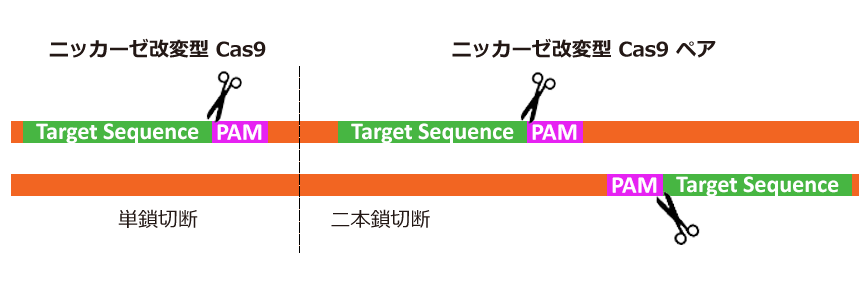
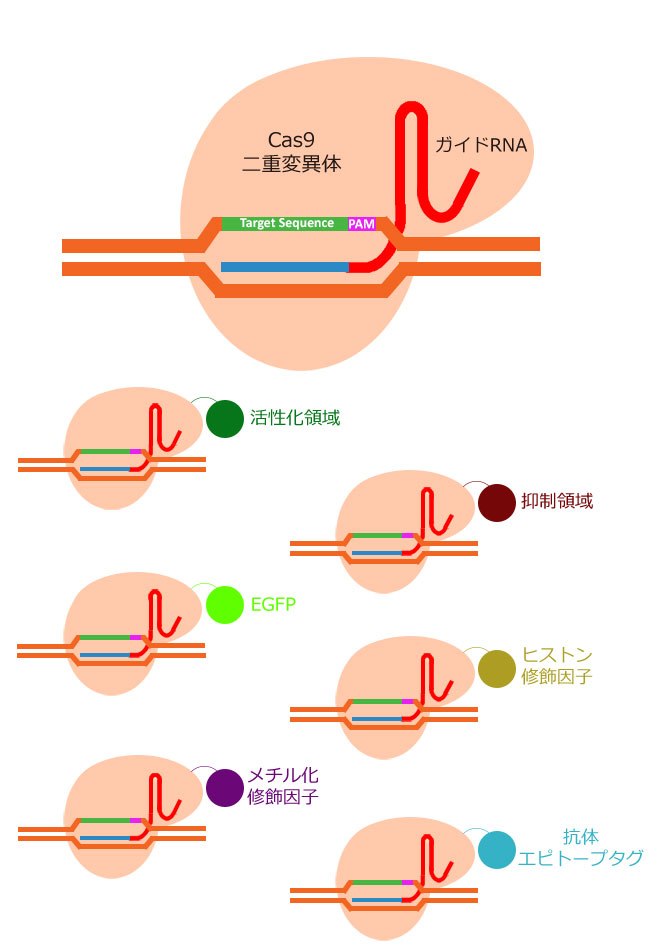
引用元:CRISPR-Cas system for editing, regulation and targeting genomes. Sander, Jeffry D and Joung, J Keith. 4, s.l. : Nature Biotechnology, March 2, 2014, Vol. 32
もう1つの顕著な違いは、Cas9 は切断後に平滑末端を生成するのに対し、Cpf1は方向性クローニングに使用できる5' オーバーハングの付着末端を残すことです。制限酵素などによる付着末端の生成では、Cpf1よりも認識配列が短いため、特異性が低くなります。Cpf1のこの特性は、in vitro で非常に特異的な DNAアセンブリを実行するために利用されています (28)。この方法を in vivo で使用すると、神経などの非分裂細胞にDNAノックインが可能です。この場合、HDRを介したゲノム編集は特に困難です。
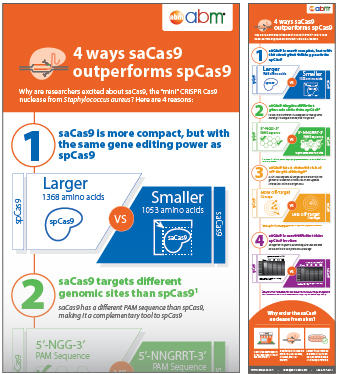
S. pyogenes Cas9(spCas9)は最も一般的に使用されるCRISPRヌクレアーゼですが、最近注目されているのは、黄色ブドウ球菌(saCas9)から分離された小型Cas9ヌクレアーゼです。この小さなヌクレアーゼは、生物におけるCRISPRベースの遺伝子編集に携わる生物医学研究業界を変える大きな可能性を秘めています。saCas9 と spCas9は、同等の効率で in vivoで真核生物のDNAを切断することができます(32)。 ただし、saCas9にはいくつかの特性があり、特定のアプリケーションでより便利に使用できます。
saCas9 は in vitro で spCas9よりも効率的:spCas9と比較して、saCas9は DNA切断に必要な酵素と時間が少なくて済みます。
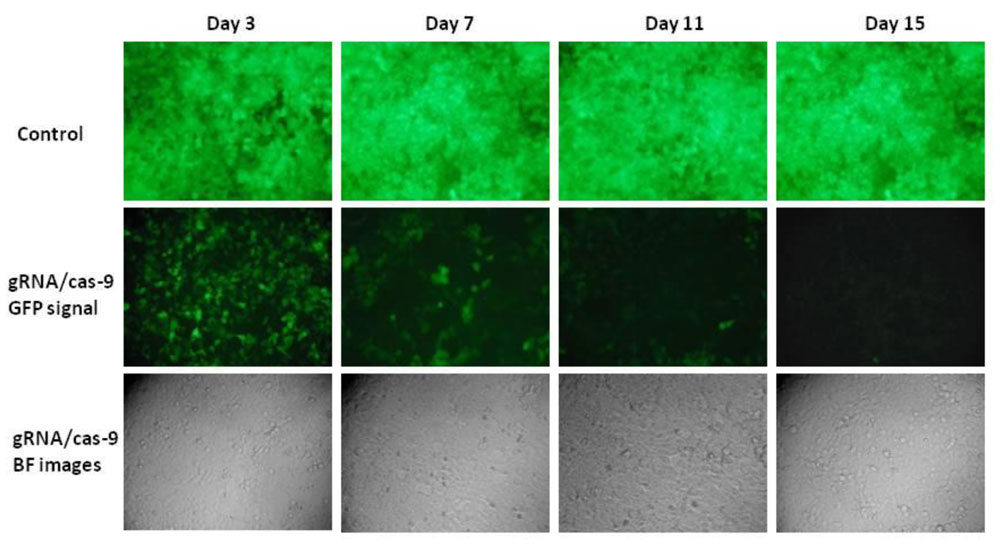
図9. S. pyogenes Cas9(spCas9)はCRISPR遺伝子編集でよく知られているヌクレアーゼですが、黄色ブドウ球菌(saCas9)からミニチュアCas9ヌクレアーゼが追加されたことで、多くの研究者が in vivo で遺伝子編集機能を拡張できる可能性に期待を寄せています。spCas9と同様に、saCas9 は二本鎖DNAを切断することができますが、spCas9とは異なり、saCas9の特色であるサイズ、PAMシーケンスなどにより、spCas9よりも優位性があります。
引用元:CRISPR/Cas9 for genome editing: progress, implications and challenges. Zhang, Feng, Wen, Yan and Guo, Xiong. 1, s.l. : Human Molecular Genetics, March 17, 2014, Vol. 23.; CRISPR-Cas system for editing, regulation and targeting genomes. Sander, Jeffry D and Joung, J Keith. 4, s.l. : Nature Biotechnology, March 2, 2014, Vol. 32.; and ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Gaj, Thomas, Gersbach, Charles A and Barbas III, Carlos F. 7, s.l. : Cell, July 2013, Vol. 31, pp. 397-406.
Cas9–crRNA ribonucleoprotein complex mediates specific DNA cleavage for adaptive immunity in bacteria. Gasiunas, Giedrius, et al. s.l. : PNAS, September 4, 2012, Vol. 109, pp. E2579-E2586.
The CRISPRdb database and tools to display CRISPRs and to generate dictionaries of spacers and repeats. Grissa, Ibtissem, Vergnaud, Gilles and Pourcel, Christine. s.l. : BMC Bioinformatics, May 23, 2007, Vol. 8, p. 172.
CRISPR/Cas9 for genome editing: progress, implications and challenges. Zhang, Feng, Wen, Yan and Guo, Xiong. 1, s.l. : Human Molecular Genetics, March 17, 2014, Vol. 23.
A Guild of 45 CRISPR-Associated (Cas) Protein Families and Multiple CRISPR/Cas Subtypes Exist in Prokaryotic Genomes. Haft, Daniel H, et al. 6, November 11, 2005, PLoS Computational Biology, Vol. 1, pp. 474-483.
Evolution and classification of the CRISPR–Cas systems. Makarova, Kira S, et al. s.l. : Nature Reviews Microbiology, Jun 2011, Vol. 9, pp. 467-477.
A Programmable Dual-RNA–Guided DNA Endonuclease in Adaptive Bacterial Immunity. Jinek, Martin, et al. 6096, Jun 28, 2012, Science, Vol. 337, pp. 816-821.
CRISPR-Cas system for editing, regulation and targeting genomes. Sander, Jeffry D and Joung, J Keith. 4, s.l. : Nature Biotechnology, March 2, 2014, Vol. 32.
Donor DNA Utilization During Gene Targeting with Zinc-Finger Nucleases. Beurner, Kelly J, et al. 4, s.l. : Genes Genomes Genetics, March 11, 2013, Vol. 3, pp. 657-664.
RNA-guided editing of bacterial genomes using CRISPR-Cas systems. Jiang, Wenyan, et al. s.l. : Nature Biotechnology, January 29, 2013, Vol. 31, pp. 233-239.
Multiplex Genome Engineering Using CRISPR/Cas Systems. Cong, Le, et al. 6121, s.l. : Science, January 3, 2013, Vol. 339, pp. 819-823.
Interference by clustered regularly interspaced short palindromic repeat (CRISPR) RNA is governed by a seed sequence. Semenova, Ekaterina, et al. 25, s.l. : PNAS, Jun 21, 2011, Vol. 108, pp. 10098-10103.
RNA-guided complex from a bacterial immune system enhances target recognition through seed sequence interactions. Wiedenheft, Blake, et al. 25, s.l. : PNAS, June 21, 2011, Vol. 108.
High-frequency off-target mutagenesis induced by CRISPR-Cas nucleases in human cells. Fu, Yanfang, et al. s.l. : Nature Biotechnology, June 23, 2013, Vol. 31, pp. 822-826.
Improving CRISPR-Cas nuclease specificity using truncated guide RNAs. Fu, Yanfang, et al. s.l. : Nature Biotechnology, January 26, 2014, Vol. 32, pp. 279-287.
CRISPR RNA–guided activation of endogenous human genes. Maeder, Morgan, et al. s.l. : Nature Methods, July 25, 2013, Vol. 10, pp. 977-979.
RNA-guided gene activation by CRISPR-Cas9–based transcription factors. Perez-Pinera, Pablo, et al. s.l. : Nature Methods, July 25, 2013, Vol. 10, pp. 973-976.
CRISPR-Mediated Modular RNA-Guided Regulation of Transcription in Eukaryotes. Gilbert, Luke A, et al. 2, s.l. : Cell, July 11, 2013, Vol. 154, pp. 442-451.
Locus-specific editing of histone modifications at endogenous enhancers. Mendenhall, Eric M, et al. s.l. : Nature Biotechnology, September 08, 2013, Vol. 31, pp. 1133-1136.
Cpf1 Is A Single RNA-Guided Endonuclease Of A Class 2 CRISPR-Cas System. Zetsche, B. et al. 2015, Cell, Vol. 163, pp. 759-771.
Genome-wide specificities of CRISPR-Cas Cpf1 nucleases in human cells. Kleinstiver, BP. et al. 2016, Nat. Biotechnol., Vol. 34, pp. 869-874.
Advances In Genomic Sequence Analysis And Pattern Discovery. Elnitski, L., Piontkivska, H., & Welch, LR. Science, Engineering, and Biology Informatics, 2011, World Scientific.
CRISPR-Cpf1 assisted genome editing of Corynebacterium glutamicum. Jiang, Y. et al. 2017. Nat. Commun., Vol. 8, 15179.
CRISPR/Cas9 mediated targeted mutagenesis of the fast growing cyanobacterium Synechococcus elongatus UTEX 2973.Wendt, KE. et al. 2016, Microb. Cell Fact. Vol. 15, 115.
The CRISPR-associated DNA-cleaving enzyme Cpf1 also processes precursor CRISPR RNA. Fonfara, I., et al. 2016, Nature, Vol. 532, pp. 517-521.
Chemically Modified Guide RNAs Enhance CRISPR-Cas Genome Editing In Human Primary Cells. Hendel, A. et al. 2015, Nat. Biotechnol. Vol. 33, 985-989.
Multiplex gene editing by CRISPR-Cpf1 using a single crRNA array.Zetsche, B. et al. 2017, Nat. Biotechnol., Vol. 35, pp. 31-34.
The CCTL (Cpf1-assisted Cutting and Taq DNA ligase-assisted Ligation) method for efficient editing of large DNA constructs in vitro. Lei, C. et al. Nucleic Acids Res., Vol. 45, e74.
Genome Engineering Using The CRISPR-Cas9 System. Ran, FA. et al. 2013, Nat. Protoc., Vol. 8, pp. 2281-2308.
CRISPR-Cpf1 correction of muscular dystrophy mutations in human cardiomyocytes and mice. Zhang, Y. et al. 2017, Science Advances, Vol. 3, e1602814.
CRISPR/Cpf1-mediated DNA-free plant genome editing.Kim H. et al. 2017, Nat. Commun., Vol. 8, 14406..
Efficient Production of Gene-Modified Mice using Staphylococcus aureus Cas9. Zhang X. et al. 2016, Sci. Rep., Vol. 6, 32565.
In Vivo Genome Editing Using Staphylococcus Aureus Cas9. Ran, FA. et al. 2015, Nature, Vol. 520, pp.186-191.
Broadening the targeting range of Staphylococcus aureus CRISPR-Cas9 by modifying PAM recognition. Kleinstiver, BP. et al. 2015, Nat. Biotechnol., Vol. 33, pp.1293-1298.
Dynamic Imaging of Genomic Loci in Living Human Cells by an Optimized CRISPR/Cas System. Chen, Baohui, et al. 7, s.l. : Cell, December 19, 2013, Vol. 155, pp. 1479-1491.
ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Gaj, Thomas, Gersbach, Charles A and Barbas III, Carlos F. 7, s.l. : Cell, July 2013, Vol. 31, pp. 397-406.
A Programmable Dual-RNA-Guided DNA Endonuclease in Adaptive Bacterial Immunity. Jinek, Martin, et al. s.l. : Science, June 28, 2012, Vol. 17, pp. 816-821.
Genome engineering in Saccharomyces cerevisiae using CRISPR-Cas systems. DiCarlo, James E, et al. s.l. : Nucleic Acids Research, April 12, 2013, Vol. 10, pp. 1-8.
Multiplex and homologous recombination-mediated genome editing in Arabidopsis and Nicotiana benthamiana using guide RNA and Cas9. Li, Jian-Feng, et al. s.l. : Nature Biotechnology, August 08, 2013, Vol. 31, pp. 688-691.
Efficient genome editing in zebrafish using a CRISPR-Cas system. Hwang, Woong Y, et al. s.l. : Nature Biotechnology, January 29, 2013, Vol. 31, pp. 227-229.
Highly Efficient Genome Modifications Mediated by CRISPR/Cas9 in Drosophila. Yu, Zhongsheng, et al. s.l. : Genetics, July 5, 2013, Vol. Early Online.
Heritable genome editing in C. elegans via a CRISPR-Cas9 system. Friedland, Ari E, et al. s.l. : Nature Methods, June 30, 2013, Vol. 10, pp. 741-743.
One-Step Generation of Mice Carrying Mutations in Multiple Genes by CRISPR/Cas-Mediated Genome Engineering. wang, Haoyi, et al. 4, s.l. : Cell, May 9, 2013, Vol. 153, pp. 910-918.
Harnessing the CRISPR/Cas9 system to disrupt latent HIV-1 provirus. Ebina, Hirotaka, et al. s.l. : Scientific Reports, August 26, 2013, Vol. 3, p. 2510.
Protospacer Recognition Motifs - Mixed Identities and Functional Diversity. 5, s.l. : RNA Biology, May 2013, Vol. 10, pp. 891-899.























 このページを印刷する
このページを印刷する