





















 このページを印刷する
このページを印刷する
広島大学大学院統合生命科学研究科 教授
ゲノム編集イノベーションセンター長
山本 卓 先生
ゲノム編集イノベーションセンター長
山本 卓 先生
![]()

ゲノム編集は、人工DNA切断酵素(ゲノム編集ツール)を用いて標的遺伝子を自在に改変することが可能な新しい技術です。微生物から動物や植物まで様々な生物での遺伝子改変が可能なことから、ゲノム編集はライフサイエンス研究において必要不可欠な技術となりました。これは、2012年に発表された第三世代のゲノム編集ツールCRISPR-Cas9によって実現し、簡便かつ効率的なゲノム編集は、この10年間で全ての研究者の技術になったと言っても過言ではありません。CRISPR-Cas9のように開発スピードの速い技術は近年例がなく、基礎研究のみならず応用研究での利用も積極的に進められています。本稿では、ゲノム編集ツール開発の歴史、ゲノム編集を用いた標的遺伝子改変、ゲノム編集の発展技術について紹介します。
ゲノム編集技術の基盤となるゲノム編集ツールは、人工制限酵素(人工ヌクレアーゼ)とRNA誘導型ヌクレアーゼに大別されます1)。人工ヌクレアーゼとして開発されたZinc-finger nuclease(ZFN)とTALEN(Transcription activator-like effector nuclease)は、目的の塩基配列に特異的に結合するDNA認識・結合ドメインに制限酵素FokIのDNA切断ドメインを連結した人工ヌクレアーゼで(図1)、一組で発現させることによって目的の塩基配列にDNA二本鎖切断(DSB : double-strand break)を誘導することができます。ZFNはジンクフィンガーの連結体をTALENはTALEリピートと呼ばれる繰り返しモチーフをDNA認識・結合ドメインとして利用しています。最近、特異性と切断活性の高い国産のPlatinum TALENが開発され、産業分野での活用も進められています。
RNA誘導型ヌクレアーゼであるCRISPR(Clusteredregularly interspaced short palindromic repeats)-Cas9(CRISPR-associated protein 9)は、2012年にScience誌に初めて報告されました2)。人工ヌクレアーゼが、標的配列への結合にDNA結合ドメインを利用するのに対して、CRISPR-Cas9は、短鎖RNAと標的DNAの塩基対形成を利用します。真正細菌や古細菌は、切断したファージDNAをCRISPR領域に取り込み、細胞に記憶します。再びファージが侵入すると、取り込んだDNAを鋳型とした短鎖のcrRNAを合成します。さらに、crRNAはtracrRNAとCas9ヌクレアーゼとの複合体を形成し、PAM(Protospacer AdjacentMotif)を目印にして塩基対形成を介して標的配列を切断します。ゲノム編集では、crRNAとtracrRNAを1分子のsgRNA(single guide RNA)として作製し、sgRNAとCas9の2因子の発現(あるいはCas9タンパク質とsgRNAの複合体の導入)によって内在の遺伝子破壊が可能です(図1)。現在多くの研究者が、化膿レンサ球菌(S. pyogenes)のSpCas9を用いており、PAMとして利用できる3塩基(5’-NGG-3’)があれば基本的に標的遺伝子の改変が可能です。CRISPRCasの利点は、sgRNAの種類を増やすことによって、多重遺伝子改変が簡単に行えることで、培養細胞でも生物個体でも多数の成功例が既に報告されています。
CRISPRでは、類似配列への変異導入(オフターゲット作用)について注意が必要です1)。標的遺伝子の遺伝子破壊が成功する一方で、類似する配列に欠失や挿入などの変異を同時に導入する可能性です。オフターゲット作用の程度は、ゲノム編集ツールの種類や用いる細胞や個体によって異なりますが、特にガン細胞では高い傾向にあります。そのため、PAMの制約を回避する開発や特異性をあげる研究が、立体構造解析を基盤として進められています。PAMの制約を回避したCRISPRとしてはSpCas9-NG(PAMは5’-NG-3’)が開発されています3)。また、Cas9のアミノ酸配列を改変することによってオフターゲット作用を低減した複数の変異体が開発されています。eSpCas9(enhanced specificity Cas9)やHiFiCas9(Cas9 high-fidelity variant)では、高い切断活性を維持しつつオフターゲット作用を低減する改変に成功しています4)、5)。
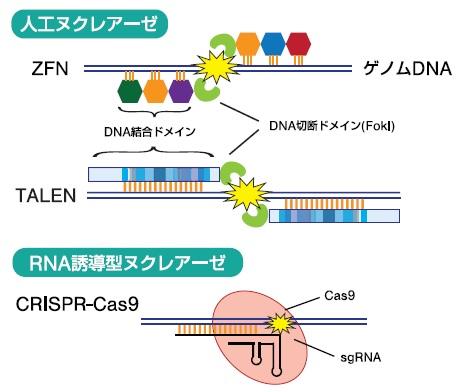
図1 各ゲノム編集ツールの概要
ゲノム編集ツールによって導入されたDSBは、細胞内の複数のDSB修復経路によって修復されます。主に、非相同末端連結(NHEJ : non-homologous end-joining)と相同配列依存的修復(HDR : homology-directed repair)の2つの経路によって修復されますが、繰り返しDSBを導入することによって修復エラーを誘導し、標的遺伝子の改変を行います。NHEJ経路では、修復過程のエラーによって、欠失や挿入などの変異を導入することが可能です。これによって、遺伝子内の変異であれば、フレームシフトを引き起こし遺伝子が破壊されます(遺伝子ノックアウト、(図2))。また、HDR経路の修復では、ドナーベクターを共導入することによって、切断部分に外来の遺伝子を挿入することが可能です(遺伝子ノックイン、(図2))。最近では、ドナーベクターの代わりに短鎖の一本鎖DNA(ssODN)を利用した一塩基レベルのノックインも盛んに行われています。
NHEJ経路を利用した遺伝子ノックアウトは、基本的に全てのゲノム編集ツールで可能であり、多くの成功例が報告されています。最近では、微生物などこれまで標的遺伝子改変が難しかった生物種での遺伝子ノックアウトの成功例も増えてきています。一方、HDR経路の中でも相同組換え(HR ;homologous recombination)を利用した遺伝子ノックインは、細胞種や生物種によって効率が異なるので注意が必要です。HRの活性は、細胞周期に依存しており、全くノックインできないケースも見られます。また、ゲノム編集の効率は、細胞であれば遺伝子導入効率、個体であればマイクロインジェクションが可能かどうかなど実験手法によっても影響を受けます。これらの問題を解決する方法として、NHEJを利用した新しい遺伝子ノックイン法(HITI法)が最近開発されました6)。この方法では、NHEJによって切断された箇所にレポーター遺伝子を挿入されますが、両方向に挿入された遺伝子のうち、目的と反対の方向に入った場合は再切断を誘導するように工夫されています。これによって一方向の正確なノックインアレルが作られます。また、筆者のグループは、HDRの一つである短いホモロジー配列(マイクロホモロジー)を介したMMEJ修復によって簡便にノックインする方法(PITCh法)を開発しており、現在では、研究目的に合った遺伝子ノックイン法の選択が可能となっています7)。ゲノム編集技術では、同時に複数箇所を切断することによって、染色体レベルでの編集も可能であることも報告されています。同一染色体上の2箇所を切断すると大規模な欠失を作製することが培養細胞や個体のレベルで証明されています。欠失と比較すると頻度は低いですが、同一染色体上の2箇所の切断によって逆位を誘導することも可能となっています。これらの染色レベルの実験を行う場合、CRISPR-Cas9であれば複数のsgRNAを使うだけで実行可能です。筆者は、最近一つのベクター中に7種類のsgRNAの発現カセットを組み込んだオールインワンベクターを開発しており8)、このベクターを用いることによって効率的に染色体レベルの改変が可能になると考えています。
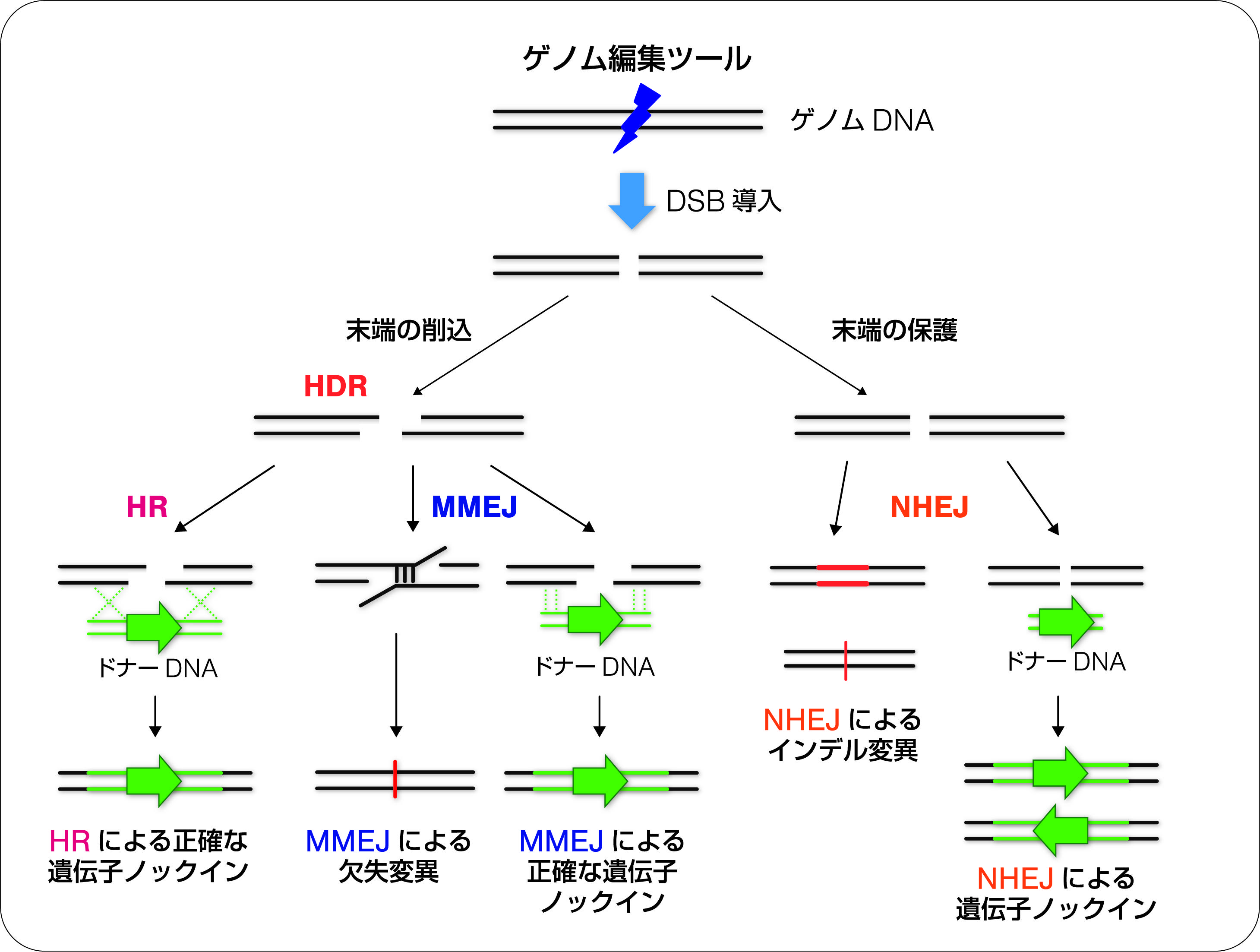
図2 ゲノム編集ツールによる遺伝子改変の概略
ゲノム編集では、任意の塩基配列を認識・結合するシステムとDNA切断ドメインを連結したゲノム編集ツールが基盤となりますが、このDNA切断ドメインの替わりに様々なタンパク質の機能ドメイン連結した発展技術が開発されています(図3)。例えば、転写調節因子の転写活性化(あるいは転写抑制)ドメインを連結した人工の転写活性化(転写抑制)因子によって、内在の遺伝子発現の転写量を増大させることに成功しています。また、DNAやヒストンの修飾酵素の機能ドメインを連結させたエピゲノム編集因子は、標的の遺伝子発現する新しい技術と期待されています。
脱アミノ化酵素(デアミナーゼ)を連結させた塩基編集技術(べースエディティング)は、DSBの導入を回避して一塩基改変が可能です9)。CからTあるいはAからGへの改変酵素が開発され疾患SNPの再現や修復がさらに最近、逆転写酵素を連結したプライムエディティングは、pegRNAとよばれるガイドRNA中に目的の変異を導入することによって、ドます。プライムエディティングによって、培養細胞や動植物個体での塩基編集の成功例が既に報告されています10)。
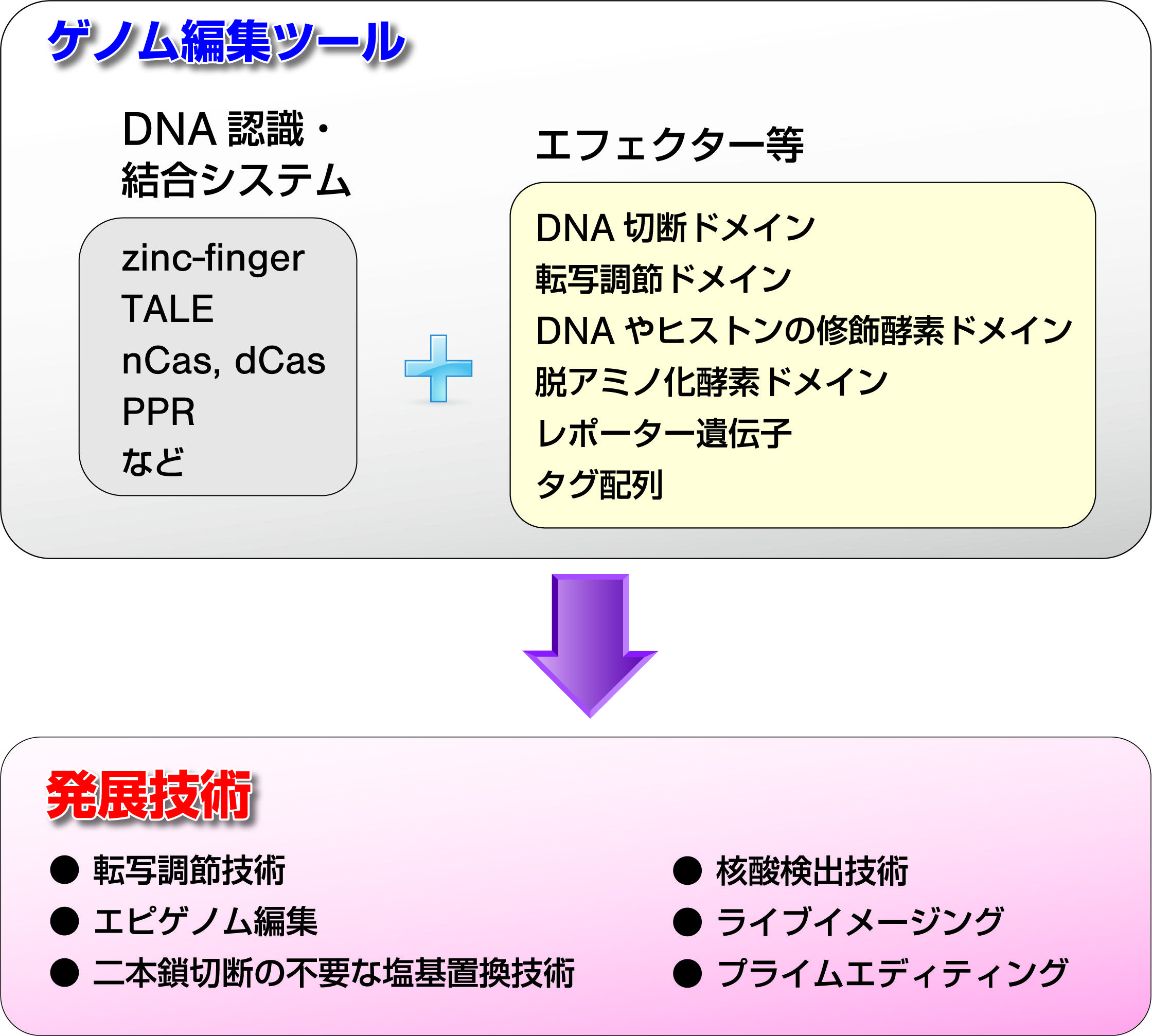
図3 様々なゲノム編集ツールと発展技術
ゲノム編集は、様々な生物での遺伝子ノックアウトと遺伝子ノックインを基盤に、高度な改変技術の開発や応用研究へ展開していくと予想されます。これらの研究開発によって、これまで理解が困難であった生命現象の解明や産業への適用が、アイディア次第で可能になるでしょう。CRISPRでは、CRISPR-Cas9を代表とするエフェクター因子が単一のクラス2が開発の主流ですが、複数のエフェクター因子とsgRNAで標的を切断するクラス1が国産技術として開発されました11)、12)。これらクラス1のCRISPRはsgRNAが長いため、特異性を高めたゲノム編集が可能です。切断によって大きな欠失を導入するという共通の性質も見られます。国内ではこれらのゲノム編集技術を積極的に利用し、基礎開発と応用開発を深化させていくことが重要だと考えています。日本ゲノム編集学会では、初心者向けの講習会を行っており、この講習会を利用してゲノム編集研究を始めるきっかけにして頂ければと願っています。
商品は「研究用試薬」です。人や動物の医療用・臨床診断用・食品用としては使用しないように、十分ご注意ください。
※ 表示価格について

お問い合わせ
商品に関するご相談・お問い合わせ
![]()
受付時間:平日 (9:00〜17:30)








© COSMO BIO





