






















 このページを印刷する
このページを印刷するミニ特集 細胞内シグナル伝達研究のトピックス 総説:GTP結合タンパク質 〜細胞の増殖・分化など様々な生命現象の制御〜
- カテゴリから探す > シグナル伝達 > リン酸化/ユビキチン化/SUMO化 > GTP結合タンパク質
- カテゴリから探す > シグナル伝達 > シグナル伝達
- カテゴリから探す > 特集 > 研究ターゲット
総説
◆バイオタイマーとしてのGTP結合タンパク質
高等生物は、外界及び自らの体内の様々な情報をとらえ、適切に対処し、自らの恒常性を維持しています。この恒常性維持のために、生物は神経系と、内分泌系の2つの大きな情報伝達系が大きな役割をしており、この情報伝達機能が正常に働かないと、生体の恒常性が崩れ、様々な病気を引き起こすことになります。神経系と内分泌系は多くの神経伝達物質やホルモンなどで細胞機能を調節しますが、この調節機能の多くの系で細胞内においてGTP結合タンパク質(Gタンパク質)を介した経路で情報伝達が行われています。Gタンパク質は、大きく分けて三量体Gタンパク質と、低分子量Gタンパク質の2つがあります。三量体Gタンパク質は、α、β、γの3つのヘテロコンプレックスで、分子量、約40kDaのαサブユニットがGTPase活性を有します。一方、低分子量Gタンパク質は分子量、約20kDa程度の単分子のGTPaseです。Gタンパク質は、GDP結合型が不活性型で、受容体などからの刺激を受けてGTP結合型となり、活性化され、情報を伝達します。このように、Gタンパク質はGDP結合不活性型とGTP結合活性型の間をサイクル(GTPaseサイクル)することにより、細胞外からの情報を細胞内に伝達しますが、この活性調節が時間的、空間的に制御されており、バイオタイマーとしての働きをしています。
◆三量体Gタンパク質
図1に示すように三量体Gタンパク質は、そのαサブユニットの相同性と機能をもとに4つの大きなグループ(Gαs、Gαi、Gαq、Gα12)に分かれます。GαsはGαsとGαolfからなり、Gαsには選択的RNAスプライシングにより複数のアイソフォームが存在します。Gαolfは嗅細胞に特異的に発現しているGタンパク質です。GαsとGαolfは共にアデニル酸シクラーゼを活性化します。Gαiグループにはさらに4つのサブグループが存在し、Gαi、Gαo、Gαt、Gαzに分かれます。GαiはGαi1 、Gαi2 、Gαi3よりなり、Gαi1は主に脳に発現しています。エフェクターは3種類ともアデニル酸シクラーゼの抑制であり、機能的な差異はありません。Giに共役する受容体はMAPキナーゼを活性化し、細胞増殖を促進することが知られていました。この機能発現に関わるGαiのエフェクターとして、低分子量Gタンパク質、RapのGTPase活性を促進し、Rapの活性を抑制するRap1GAPIIが同定されました。RapはRasのエフェクターであるRafに結合し、Ras-RafによるMAPキナーゼの活性化を抑制します。活性型GαiはRap1GAPIIに結合し、細胞膜にリクルートし、Rapの活性を低下させることにより、RapによるRas-Rafの抑制を解除することにより、結果としてMAPキナーゼを活性化します。GαoにはGαo1、Gαo2の2種類があり、脳、神経系に局在しています。In vitroではGαiと同様にアデニル酸シクラーゼの抑制活性を示しますが、生理的な神経機能はいまだ不明です。Gαoは神経軸索の先端の成長円錐に強く発現していることがわかっています。最近、Gαoの新規のエフェクターとしてGRIN1が見出され、神経突起形成に関わるのではないかと推定されています。Gαtには2種類のサブタイプ、Gαt1、Gαt2があり、それぞれ網膜の桿体、錐体に発現し、ロドプシン、オプシンにより活性化され、cGMPホスホジエステラーゼの活性を抑制し、光の情報伝達に関わっています。GαzはGαiと機能は全く同じですが、C末端から4番目にある、百日咳毒素によりADPリボシル化されるシステイン残基がなく、百日咳毒素に対する感受性で異なります。Gα12ファミリーは、三量体Gタンパク質の中で最後に見つかったサブファミリーで、 ほ乳類ではGα12とGα13の2つからなります。現在までに、いくつかのエフェクター分子が同定されていますが、Gα12とGα13は共に共通のエフェクターを介して同じシグナルを流すものと考えられています。しかし、Gα12とGα13それぞれの遺伝子を欠損したノックアウトマウスが作成されており、Gα12が致死であるのに対し、Gα13は生まれてくることから、Gα12とGα13は異なる生理機能を担っているものと考えられています。Gα12ファミリーの最もよく知られているエフェクターは、低分子量Gタンパク質、Rhoを活性化するRGS-RhoGEFです。RGS-RhoGEFには3種類、p115、PZD-RhoGEF、LARG、が知られており、p115は血球系に主要に発現しています。3種類ともGα12に対しGAP活性を示すRGSドメインを持ち、このドメインにGα12ファミリーが結合し、RhoGEF活性を促進します。その結果、Rhoを活性化することによって、ストレスファイバー形成や神経突起退縮などの様々なRhoの生理機能を発現させます。Gα12ファミリーはRho活性化以外にカドヘリンやラディキシンに結合することが示されましたが、その生理的意義は不明です。我々は、Gα12ファミリーのエフェクターとして、セリン・スレオニン型プロテインホスファターゼ5(PP5)を同定しました。PP5は他のホスファターゼと異なり、basal活性が低く、G α1 2ファミリーがPP5のN末端側にあるTPR (tetratricopeptide repeat)ドメインに結合すると、TPRドメインによる触媒部位への抑制が解除されて、活性化されます。三量体Gタンパク質のエフェクターの中でPP5は、ホスファターゼとして初めての例です。
三量体Gタンパク質は一部の例外を除いて細胞膜を7回貫通する構造をしたロドプシン型の受容体により活性化されます。受容体と三量体Gタンパク質との共役の特異性は決まっており、βアドレナリン受容体はGsを、α1アドレナリン受容体はGqと特異的に共役し、細胞内での情報伝達の特性が決まっています。しかし、プロスタグランジンEP3受容体のように、GiとG12の2つのGタンパク質と共役する受容体も多く存在します。受容体には三量体Gタンパク質のαサブユニットのGDP-GTP交換反応を促進し、αサブユニットをβγサブユニットから乖離(かいり)させ、GTP結合αサブユニットによるエフェクターの活性化を促進します。受容体にはGタンパク質活性化ドメインがあり、受容体により異なりますが、第2細胞内ループや第3細胞内ループに存在します。事実、このドメインの合成ペプチドにより、直接Gタンパク質を活性化できます。しかし、受容体はリガンドが結合しなければGタンパク質を活性化しません。これは、受容体内のGタンパク質活性化ドメインによる活性化を抑制する機構があるためで、C末端鎖がこの抑制に関わっている場合が見出されています。そして、リガンドが結合すると、この抑制が解除されて、活性化ドメインによる活性化が起こります。
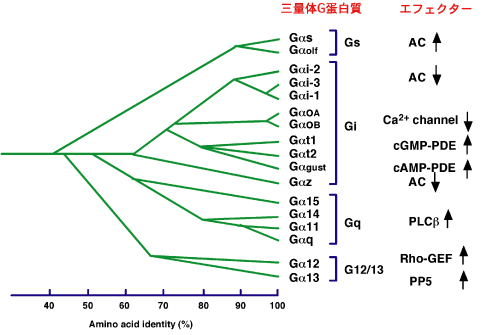
◆図1.三量体Gタンパク質の種類とエフェクター
AC:アデニル酸シクラーゼ, PDE:ホスホジエステラーゼ, PLC:ホスホリパーゼC,
PP5:セリン・スレオニンプロテインホスファターゼ5
◆低分子量Gタンパク質
低分子量Gタンパク質は、Rasスーパーファミリーと称されており、さらに、Ras、Rho、Rab、Arf、Ranの5つのサブファミリーに分かれます。Rasファミリーには代表的なRasであるH-Ras、N-Ras、K-Ras以外にRapやRalサブファミリーなど様々な種類があり、細胞の増殖や分化に重要な役割を果たしています。RhoファミリーにはRho、Rac、Cdc42をはじめほ乳類で22種類が報告されており、細胞骨格の制御を通して細胞の形態調節に関わるGタンパク質です。Rabは30種類以上存在し、小胞輸送に関与します。Arfは6種類あり、やはり小胞輸送に関わっています。Ranは核内への物質輸送に関与する分子です。 低分子量Gタンパク質は、三量体Gタンパク質と同様にGTP結合活性型とGDP結合不活性型の間でGTPaseサイクルを行います。通常、Gタンパク質はGDP-GTP交換因子(GEF; guanine nucleotide exchange factor)により活性型となり、それぞれに特異的なエフェクターに結合し、生理作用を発揮します。また、GTPase活性化因子(GAP;GTPase activating protein)により内在性のGTPase活性が促進され、GTPを水解して不活性型となります。従って、低分子量Gタンパク質はGEFとGAPによりその活性が制御され、細胞外からの様々なシグナルはGEFかGAPを活性化することによりGタンパク質の活性を調節しています。低分子量Gタンパク質はそのほとんどでC末端から4番目のアミノ酸残基がシステインであり、ファルネシル基やジェラニルジェラニル基などの脂肪酸でファルネシル化されており、この脂質修飾により、細胞膜に直接結合できると考えられています。RhoとRabファミリーに関して、GDP結合不活性型の時はそれを特異的に認識し結合するG D I( G D Pdissociation inhibitor)と複合体を形成し、細胞質に局在します。この結合はGDIがGタンパク質のファルネシル基を認識して細胞膜への結合ができないようにしています。GDIの中で、RhoGDI3はRhoGを認識しますが、細胞質ではなく、ゴルジ体に局在します。一方、Rasファミリーに結合するGDIはなく、上述した脂質修飾により常に細胞膜に結合していると考えられています。
Rasスーパーファミリー全ての機能を述べるスペースはないので、ここではRhoファミリーを代表として解説します。代表的なRhoファミリーで非常によく研究されているのはRhoA、Rac、Cdc42の3種類です。これらRhoファミリーの機能は繊維芽細胞など培養細胞のアクチン骨格に対する作用の研究を発端として始まりました。そして、RhoAはストレスファイバー、Racはラメリポディア、Cdc42はフィロポディア形成を引き起こします。それぞれのGタンパク質の機能発現に関わる多くの特異的なエフェクターがクローニングされ、その機能が解析されています。RhoAの代表的なエフェクターはRhoキナーゼです。Rhoキナーゼは、ミオシンホスファターゼのサブユニットMBS(myosin binding subunit)をリン酸化することにより、ホスファターゼの活性を抑制すると共に、ミオシン軽鎖を直接リン酸化することにより、ミオシンを活性化し、アクトミオシンの収縮を引き起こします。その結果、ストレスファイバーが形成されます。Rhoキナーゼはミオシンの活性化により、神経突起の退縮や細胞分裂にも重要な役割を果たしています。RhoAの他のエフェクターとして、mDiaがあり、アクチン重合を調節しています。一方、Racのエフェクターも多くが同定されており、その中で、IRSp53やNap-PIR121-Abi2に結合することにより、WAVEを介してアクチン重合を促進し、ラメリポディアの形成を引き起こします。Cdc42のエフェクターはN-WASPであり、WAVEと同様にアクチン重合を促進し、フィロポディア形成を引き起こします。細胞の遊走において、細胞の前進方向にある先導端で、RacとCdc42はWAVEやN-WASPによりラメリポディアやフィロポディア形成により、ダイナミックに細胞膜の伸展を推進する一方で、細胞の後方部でRhoARhoキナーゼによるアクトミオシンの収縮作用により細胞後部の牽引を起こすことにより、細胞は前進します。このように、上述したRhoA、Rac、Cdc42とそれらのエフェクターの連携した働きが速やかな細胞運動を推進することになります。Rhoファミリーの活性化因子、GEFは、大きく分けて2つのグループがあります。1つはDH-PHドメインを持つものでDH(Dbl homology)ドメインがRhoファミリーのGDPGTP交換反応を促進する触媒領域です。三量体Gタンパク質の項で述べたように、RhoAを活性化する代表的なGEFはRGS-RhoGEFで、ロドプシン型受容体によるG12ファミリーの活性化により活性が促進され、RhoAを活性化します。Racを活性化するGEFとして、Vav、Tiam、PIXなど多くの分子が同定されており、様々な細胞外からの刺激に応じて活性化されると考えられています。もう1つのグループのGEFはCDM(Ced5-Dock180-Mbc)ファミリーと呼ばれ、DHドメインを持たず、代わりに別のDHR2と呼ばれる触媒部位を持つグループです。代表的なC D M ファミリーであるDock180はN末端にSH3ドメインを持ち、ここを介してElmoという分子に結合します。Elmoは別のRhoファミリー、RhoGのエフェクターであり、RhoG-Elmo-Dock180の複合体を形成し、Racを活性化します。このRac活性化経路は、神経成長因子による神経突起伸長やインテグリンによる細胞運動促進におけるRacの活性化の重要な経路です。一方、RhoファミリーのGAPも多くの分子がクローニングされており、ほ乳類で少なくとも80種類以上あると考えられています。その1つ、p190RhoGAPはsrcなどのチロシンキナーゼによりリン酸化され、活性化されると考えられています。
Rhoファミリーには、RhoA、Rac、Cdc42以外に多くの種類があります。その中で、特にユニークなGタンパク質はRndサブファミリーであり、他のRhoファミリーと異なり、GTPase活性が極めて低く、常時活性型です。Rndサブファミリーのうち、Rnd1とRnd2は脳神経系に特異的に発現しており、Rnd1は神経軸索ガイダンス分子、Semaphorin4Dの受容体、Plexin-B1の細胞内領域に常に安定に結合しており、Plexin-B1とRhoAのGEF、PDZ-RhoGEFの結合を促進することにより、リガンドによるRhoAの活性化を起こします。その結果、細胞体退縮作用を引き起こし、軸索ガイダンス作用を調節していると考えられています。一方、Rnd2の特異的なエフェクター、Rapostlinがクローニングされ、Rnd2-Rapostlinにより、神経突起の分枝化を引き起こします。
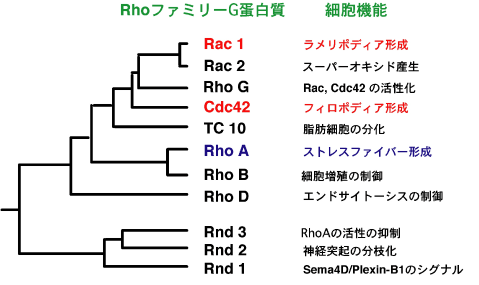
◆図2.RhoファミリーGタンパク質の種類とその代表的な機能
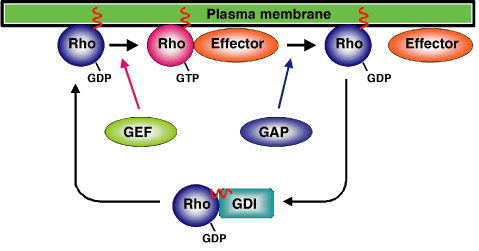
◆図3.RhoファミリーGタンパク質のGTPaseサイクル
◆Gタンパク質と疾患
以上、三量体Gタンパク質と低分子量Gタンパク質の機能について概略を述べてきました。これらGタンパク質の生体内での機能に異常が起きると様々な病気疾患が引き起こされることが知られています。偽上皮小体機能低下症では、Gαsの点変異で常時活性型となり、Gαsの分解促進によるGαsタンパク質の著しい低下が原因であることがわかりました。コレラ毒素はGαsのADPリボシル化による常時活性型による異常なcAMP産生を起こしますし、百日咳毒素はGαiのADPリボシル化による活性化阻害を引き起こします。Rasの変異による常時活性化は様々な癌を引き起こし、ヒトの癌の20%以上が何らかのRasの点変異によることが知られています。また、遺伝性精神遅滞の原因遺伝子として、RhoファミリーのGEF、GAP及びエフェクターが次々に同定され、Rhoファミリーの情報伝達が正常に行われないと、機能的な神経回路形成に異常が生ずることがわかります。このように、Gタンパク質は生体の正常な機能発現に極めて重要な役割を果たしていることがわかります。
プロフィール

根岸 学 先生
(ねぎし まなぶ)
1982年、京都大学大学院薬学研究科卒業、同年ペンシルバニア大学フィラデルフィア小児病院研究員、1985年科学技術振興事業団早石生物情報伝達プロジェクト研究員、1987年大阪バイオサイエンス研究所研究員、1990年京都大学薬学部助手、1995年同助教授、1997年同教授、1999年京都大学大学院生命科学研究科教授。研究テーマは、神経回路形成におけるGタンパク質の役割の研究。趣味は、西欧中世・ルネッサンス音楽鑑賞とそれを聴く旅。
商品は「研究用試薬」です。人や動物の医療用・臨床診断用・食品用としては使用しないように、十分ご注意ください。
※ 表示価格について

- 「総説:GTP結合タンパク質 〜細胞の増殖・分化など様々な生命現象の制御〜」は、下記のカテゴリーに属しています。
-
- カテゴリから探す > シグナル伝達 > リン酸化/ユビキチン化/SUMO化 > GTP結合タンパク質
- カテゴリから探す > シグナル伝達 > シグナル伝達
- カテゴリから探す > 特集 > 研究ターゲット













