





















 このページを印刷する
このページを印刷するHycult Biotech(ハイカルト社)の血管疾患研究における炎症と凝固解析ツールをご紹介致します。
社 製品のご紹介")
![]()

Hycult Biotech(ハイカルト社)の血管疾患研究における炎症と凝固解析ツールをご紹介致します。
背景
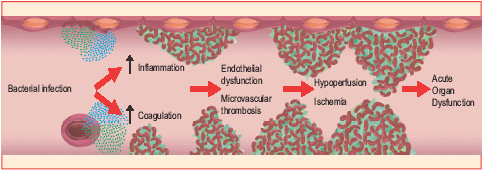
凝固系は先天性免疫系と共に発達したという証拠が増えてきています。組織損傷や微生物侵入後のシグナルパスウェイや調節回路では、驚くべきレベルの融合が起こっています。炎症メディエーターが凝固促進性シグナルを出し、血管内血栓症が先天性免疫系の複合成分を活性化します。凝固と炎症の関連性は、組み換え活性化プロテインC(APC)が敗血症治療で成功したことにより最も明確に示されています。
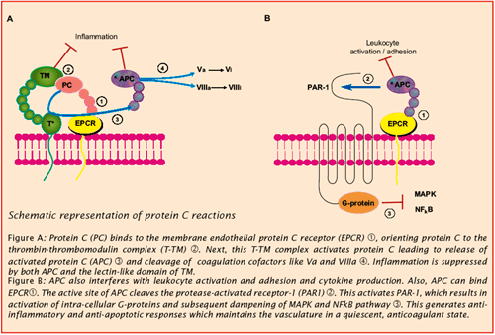
血液凝固は、出血において重要なメカニズムです。血小板プラグの形成が、血管損傷の最初の閉塞をもたらします。血液凝固は、血液の流動性維持に不可欠ないくつかの凝集素や抗凝集素により制御されています。プロテインC抗凝固パスウェイは重要な抗凝固メカニズムであり、これがまた炎症応答を制御し、炎症性サイトカインおよび虚血に応答して、場合によっては内皮細胞アポトーシスを減少させることになります。
このパスウェイの必須成分は、トロンビン、トロンボモジュリン(TM)、内皮細胞プロテインC受容体(EPCR)、プロテインC(PC)およびプロテインS(PS)です。プロテインCが、このパスウェイの重要な成分である。プロテインCは、内皮細胞膜タンパク質TMに結合したトロンビンにより活性化され、プロ酵素として循環します。いったんTMに結合すると、トロンビンは凝集促進活性を減少します。
活性化プロテインC(APC)はいくつかの凝集補足因子を開裂して抑制し、その結果、凝固系の活性をダウンレギュレーションします。APC機能は、その補足因子PSにより促進されます。また、APCは、プラスミノーゲン活性化因子抑制因子1(PAI‐1)を中和することにより線維素溶解パスウェイに影響を与えます。
プロテインCパスウェイでは、厳密には抗凝固と呼ばれるもの以外に広範囲の生物学影響を有しています。例えば、TMのレクチンドメインはNFカッパ‐ベータおよびMAPキナーゼをダウンレギュレーションする抗炎症性を有し、白血球接着および溢出を減少させます。プロテインCとAPCは、内皮表面への好中球の接着および好中球のトランスファーを直接抑制しています。
さらに、APCは、ピロリ菌感染患者の胃粘膜での炎症抑制において重要な役割をしています。APCは、虚血性脳血管構造におけるp53媒介アポトーシスをブロックすることによりこの血管構造を守っています。TMは、APCの抗炎症能力を調節しています。その際、TMは、線維素溶解の調節、細胞接着、胚発生および腫瘍成長のような生理機能も兼ね備えています。内皮細胞表面から放出された可溶性TMは血漿および尿中において検出することができ、高レベルの可溶性TMは内皮の損傷および/またはターンオーバーの促進を示します。ある種の癌では、内皮と腫瘍細胞の両者においてTMが発現しており、TM発現の喪失は予後の悪い癌と相関しています。
TMおよびEPCR両者の遺伝子転写は炎症性サイトカインによりダウンレギュレーションされるので、炎症はプロテインCパスウェイに対して重要な影響を与えます。
また、プロテインSも炎症によりダウンレギュレーションされています。血液凝固が活性化されプロテインCが消費される重篤な敗血症において、血液凝固と炎症の間におけるクロストークが十分研究されています。血漿中プロテインCレベルの低下は、微小血管血栓症の進展の一因になるものと考えられています。さらに、内皮でのTMおよびEPCRの発現レベルは、敗血症の発症期間中において減少する。APCは、敗血症と関連した有害作用を弱めます。重篤な敗血症の組み換えAPC治療の研究が、相対的死亡リスクの19%減少および6%の完全な減少に導きました。
先天性免疫系および血液凝固系は共通の先祖から始まっており、このことが2つの系の間でのクロストークを説明できます。
フィコリンは、脊椎動物による非自己認識を通じた補体媒介宿主防御に関与している蛋白質のグループであり、おそらく、無脊椎動物の先天性免疫系から脊椎動物の血液凝固系への進化における中間体でしょう。
さらに、凝固と先天性免疫の間における結合の理解および解明が、重篤な感染の研究および治療におけるさらなる理解に結びつくでしょう。
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
Anti APC, Human (Rat) Unlabeled, PC107 |
HCB | HM2151-100UG | 100 UG |
¥97,000 |
| Anti EPCR, Human (Rat) Unlabeled, RCR-252 |
HCB | HM2145-100UG | 100 UG |
¥145,000 |
| Anti EPCR, Human (Rat) Unlabeled, RCR-379 |
HCB | HM2144-100UG | 100 UG |
¥145,000 |
| Anti PAI 1, Human (Mouse) Unlabeled, MA-55F4C12 |
HCB | HM2180-100UG | 100 UG |
¥86,000 |
| Anti PAI 1, Human (Mouse) Unlabeled, MA-33H1F7 |
HCB | HM2179-100UG | 100 UG |
¥97,000 |
| Anti Protein S, Human (Rat) Unlabeled, PS7 |
HCB | HM2148-100UG | 100 UG |
¥97,000 |
| Anti CD141 / Thrombomodulin, Human (Rat) Unlabeled, RTM96 |
HCB | HM2146-100UG | 100 UG |
¥106,000 |
| Anti CD141 / Thrombomodulin, Human (Rat) Unlabeled, RTM98 |
HCB | HM2147-100UG | 100 UG |
¥106,000 |

JAM(junctional adhesion molecule)‐Aは、タイトジャンクションを通じて内皮細胞層統合性の維持に関与しているばかりでなく、単核細胞補充にも関与している。後者は、アテローム発生に対するJAM‐Aの機能的寄与を示唆しています。 JAMは約30〜40kDaの蛋白質であり、免疫グロブリンスーパーファミリーのメンバーである。JAMは、タイトジャンクションアセンブリー、白血球遊走、血小板活性化、血管形成およびウイルス結合を含む多くの細胞内プロセスにとって重要です。JAM‐A(JAM、JAM‐1またはF11 Rとしても知られている)は、内皮および上皮細胞、血小板、好中球、単球、白血球と赤血球により発現されます。JAM‐A分子の細胞外ドメインは、隣接する内皮または上皮細胞を結合させて細胞間接合、特にタイトジャンクションのまわりを安定化させる同種親和性相互作用に関与しています。JAM‐Aは、血小板凝集促進性モノクローナル抗体F11の受容体として最初に見出されました。ヒト血小板へのF11結合は、顆粒分泌、フィブリノーゲン結合および血小板凝集を引き起こしました。面白いことに、血小板減少症の患者でJAM‐Aに対する自己抗体が見出されました。 また、JAM‐Aは、白血球遊走においても重要な役割をしています。白血球遊走は、抗体により、また可溶性JAM‐A/Fc融合蛋白質により阻止することができます。しかし、白血球遊走中におけるJAM‐Aの作用についての正確なメカニズムは十分には理解されていません。白血球と内皮の間における同種親和性JAM‐A相互作用ばかりでなくβ2‐インテグリン白血球機能関連抗原‐1(LFA‐1)とのJAM‐Aの同種親和性相互作用も、白血球遊走中において活発に白血球の道案内をしているものと考えられています。ジャンクションからのJAM‐Aを分解的に誘発することにより、腫瘍壊死因子が追加的役割をしていることが示唆されます。腫瘍壊死因子(TNF)は、ジャンクションの脆弱化および内皮細胞の頂端表面へのJAM‐1再分布に導き、その結果、白血球LFA‐1との接着相互作用に利用されるようになります。 初期アテローム硬化型内皮においてJAM‐Aがアップレギュレーションされており、活性化内皮における活性化血小板の接着がJAM‐Aの同種親和性相互作用により媒介されているので、いくつかの研究はアテローム硬化症におけるJAM‐Aの役割を暗示しています。 アテローム硬化症になり易い部位において、細胞内接着分子‐1(ICAM‐1)がアップレギュレーションされており、炎症性Tリンパ球が引き寄せられています。可溶型のJAM‐AはLFA‐1のLFA‐1/ICAM‐1相互作用と拮抗し、内皮ICAM‐1に対し白血球を発現します。面白いことに、JAM‐Aは、アテローム硬化症になり易いマウスでの動脈損傷後における新生血管内膜傷害形成および単球浸潤にも関与しています。この関係において、高脂血症は、アテローム硬化型の内皮をアップレギュレーションしています。したがって、血管条件内でのJAM‐Aの役割をさらに調べ、炎症性血栓症およびアテローム発生でのJAM‐Aの役割をさらに解明することは非常に重要になるでしょう。
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
| Anti JAM-A, domain 1, Human (Rabbit) Unlabeled |
HCB | HP9041-100UG | 100 UG |
¥119,000 |
| Anti JAM-A, domain 2, Human (Rabbit) Unlabeled |
HCB | HP9042-100UG | 100 UG |
¥106,000 |
| Anti JAM-1, Human (Mouse) Unlabeled, BV16 |
HCB | HM2098-100UG | 100 UG |
¥132,000 |
| Anti JAM-1, Human (Mouse) Unlabeled, M.Ab.F11 |
HCB | HM2099-100UG | 100 UG |
¥132,000 |
| Anti JAM-1, Mouse (Rat) Unlabeled, BV12 |
HCB | HM1050-100UG | 100 UG |
¥119,000 |
| Anti JAM-C / Junctional Adhesion Molecule C, Mouse (Rat) Unlabeled, CRAM-18F26 |
HCB | HM1057-100UG | 100 UG |
販売終了 |
| Anti JAM-C / Junctional Adhesion Molecule C, Mouse (Rat) Unlabeled, 19H36 |
HCB | HM1056-100UG | 100 UG |
¥97,000 |
| Anti L-Afadin, Rat (Mouse) Unlabeled, 3 |
HCB | HM3013-100UG | 100 UG |
¥119,000 |
| Anti Nectin-2, Mouse (Rat) Unlabeled, 502-57 |
HCB | HM1052-100UG | 100 UG |
¥132,000 |
| Anti Nectin-3, Mouse (Rat) Unlabeled, 103-A1 |
HCB | HM1053-100UG | 100 UG |
¥119,000 |
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
| Anti α-V/β-3 Integrin, Human (Mouse) Unlabeled, BV3 |
HCB | HM2034-100UG | 100 UG |
¥97,000 |
| Anti Integrin αVβ3, Human (Mouse) Unlabeled, BV4 |
HCB | HM2035-100UG | 100 UG |
¥97,000 |
| Anti β-Catenin, Human (Mouse) Unlabeled, 9F2 |
HCB | HM2112-100UG | 100 UG |
販売終了 |
| Anti Integrin β1, Human (Mouse) Unlabeled, BV7 |
HCB | HM2033-100UG | 100 UG |
¥106,000 |
| Anti CD11/CD18, Human (Mouse) Unlabeled, 24 |
HCB | HM2183-100UG | 100 UG |
販売終了 |
| Anti Desmoglein-1, Human (Mouse) Unlabeled, 27B2 |
HCB | HM2113-100UG | 100 UG |
販売終了 |
| Anti Desmoglein-2, Human (Mouse) Unlabeled, 6D8 |
HCB | HM2114-100UG | 100 UG |
販売終了 |
| Anti Desmoglein-3, Human (Mouse) Unlabeled, 5G11 |
HCB | HM2115-100UG | 100 UG |
販売終了 |
| Anti E-Selectin, Human (Mouse) Unlabeled, ENA2 |
HCB | HM4003-100UG | 100 UG |
¥97,000 |
| Anti E-Selectin, Human (Mouse) Unlabeled, ENA1 |
HCB | HM4001-100UG | 100 UG |
¥97,000 |
| Anti E-selectin, Human (Rabbit) Biotin |
HCB | HP9016BT- 50UG | 50 UG |
販売終了 |
| Anti MAdCAM 1, Human (Mouse) Unlabeled, 314G8 |
HCB | HM2207-100UG | 100 UG |
¥97,000 |
| Anti CD31, Human (Mouse) Unlabeled, BV8 |
HCB | HM2039-100UG | 100 UG |
¥97,000 |
| Anti PECAM 1, Mouse (Rat) Unlabeled, MEC7.46 |
HCB | HM1013-100UG | 100 UG |
¥75,000 |
| Anti CD31, Mouse (Rat) Unlabeled, ER-MP12 |
HCB | HM1084-100UG | 100 UG |
¥75,000 |
| Anti Plakoglobin, Human (Mouse) Unlabeled, 15F11 |
HCB | HM2116-100UG | 100 UG |
販売終了 |
| Anti VCAM 1, Human (Mouse) Unlabeled, 1G11B1 |
HCB | HM4006-100UG | 100 UG |
¥75,000 |
| Anti VE Cadherin, Human (Mouse) Unlabeled, BV9 |
HCB | HM2032-100UG | 100 UG |
¥97,000 |
| Anti Vitronectin, Human (Mouse) Unlabeled, BV1 |
HCB | HM2036-100UG | 100 UG |
¥97,000 |
| Anti α-5 Integrin (Fibronectin-receptor), Human (Mouse) Unlabeled, NKI-SAM1 |
HCB | HM2126-100UG | 100 UG |
¥97,000 |
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
| Elastase ELISA kit, Human |
HCB | HK319-01 | 1*96 WELL |
¥176,000 |
| Elastase ELISA Kit, Human |
HCB | HK319-02 | 2*96 WELL |
¥285,000 |
| MPO ELISA kit, Human |
HCB | HK324-01 | 1*96 WELL |
¥162,000 |
| MPO, ELISA, Human |
HCB | HK324-02 | 2*96 WELL |
¥262,000 |
| MPO ELISA kit, Mouse |
HCB | HK210-01 | 1*96 WELL |
¥192,000 |
| MPO, ELISA, Mouse |
HCB | HK210-02 | 2*96 WELL |
¥317,000 |
| Anti Apoptotic Neutrophils, Human (Mouse) Unlabeled, BOB93 |
HCB | HM2154-100UG | 100 UG |
販売終了 |
| Anti Calprotectin, Human (Mouse) Unlabeled, 27E10 |
HCB | HM2156-100UG | 100 UG |
¥132,000 |
| Anti Galectin 3, Human (Mouse) Unlabeled, B2C10 |
HCB | HM2186-100UG | 100 UG |
¥106,000 |
| Anti MPO, Human (Mouse) Unlabeled, 266-6K1 |
HCB | HM2164-100UG | 100 UG |
¥86,000 |
| Anti MPO, Mouse (Mouse) Unlabeled, 8F4 |
HCB | HM1051-100UG | 100 UG |
¥132,000 |
| Anti MPO, Rat (Mouse) Unlabeled, 2D4 |
HCB | HM3030-100UG | 100 UG |
¥132,000 |
| Anti Neutrophils, Mouse (Rat) Unlabeled, NIMP-R14 |
HCB | HM1039-100UG | 100 UG |
¥86,000 |
| Anti Neutrophil Lipocalin [HNL, nGAL], Human (Mouse) Unlabeled, 697 |
HCB | HM2193-100UG | 100 UG |
¥119,000 |
| Anti Proteinase 3, Human (Mouse) Unlabeled, PR3G-2 |
HCB | HM2172-100UG | 100 UG |
¥97,000 |
| Anti Proteinase 3, Human (Mouse) Unlabeled, WGM2 |
HCB | HM2171-100UG | 100 UG |
¥119,000 |
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
| Anti CD36, Human (Mouse) Unlabeled, FA6-152 |
HCB | HM2122-100UG | 100 UG |
¥86,000 |
| Anti CD36, Mouse (Mouse) Unlabeled, CRF D2712 |
HCB | HM1074-100UG | 100 UG |
¥75,000 |
| Anti CD68 / Macrosialin, Mouse (Rat) Unlabeled, FA-11 |
HCB | HM1070-100UG | 100 UG |
販売終了 |
| Anti CD68, Rat (Mouse) Unlabeled, ED1 |
HCB | HM3029-100UG | 100 UG |
販売終了 |
| Anti CD163, Human (Mouse) Unlabeled, RM3/1 |
HCB | HM2157-100UG | 100 UG |
¥106,000 |
| Anti CD163, Rat (Mouse) Unlabeled, ED2 |
HCB | HM3025-100UG | 100 UG |
¥119,000 |
| Anti CD204 / SR-A, Mouse (Rat) Unlabeled, 2F8 |
HCB | HM1061-100UG | 100 UG |
販売終了 |
| Anti Dectin-1, Mouse (Rat) Unlabeled, 2A11 |
HCB | HM1067-100UG | 100 UG |
販売終了 |
| Anti Mannose Receptor, Human (Mouse) Unlabeled, 15-2 |
HCB | HM2056-100UG | 100 UG |
¥132,000 |
| Anti Mannose Receptor (CD206), Mouse (Rat) Unlabeled, MR5D3 |
HCB | HM1049-100UG | 100 UG |
販売終了 |
| Anti MARCO, Human (Mouse) Unlabeled, PLK-1 |
HCB | HM2208-100UG | 100 UG |
¥132,000 |
| Anti SR-BI, Rat (Mouse) Unlabeled, 3D12 |
HCB | HM3027-100UG | 100 UG |
¥145,000 |
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
| Anti CD34, Mouse (Rat) Unlabeled, MEC14.7 |
HCB | HM1015-100UG | 100 UG |
¥75,000 |
| Anti CD105, Human (Mouse) Unlabeled, E9 |
HCB | HM2140-100UG | 100 UG |
販売終了 |
| Anti EMAP 2, Human (Mouse) Unlabeled, 546-2 |
HCB | HM2185-100UG | 100 UG |
販売終了 |
| Anti Endostatin, Human (Mouse) Unlabeled, 1837-46 |
HCB | HM2188-100UG | 100 UG |
販売終了 |
| Anti Endothelial Cell Antigen, RECA-1, Rat (Mouse) Unlabeled, RECA-1 |
HCB | HM3012-100UG | 100 UG |
¥75,000 |
酸化ストレスおよび脂質過酸化は、アテローム発生固有の特性です。 りん脂質分子中の多価不飽和脂肪酸の触媒または非触媒酸化が、血管壁にin vivo蓄積することが知られている酸化りん脂質(OxPL)を作り出します。OxPLは好中球ではなくて単球に結合する内皮細胞の刺激のようなアテローム発生に関連した多くの生物活性を示し、それはアテローム発生の単核細胞特異性を模倣しています。OxPLは、in vitroおよびin vivoの両者でのMCP‐1,KC/IL‐8、MIP‐1アルファ、MIP‐1ベータ、RANTESおよび組織因子のようなアテローム性血栓症に関連した遺伝子の誘導を刺激します。さらに、特定のOxPLは、スカベンジャー受容体CD36に対するリガンドであり、血小板活性化因子の炎症および前凝集体効果を模倣します。OxPLは、受容体媒介または無受容体メカニズムを通じて細胞を活性化し、cAMPおよび細胞質Ca2+レベル上昇、プラテインキナーゼC,ERK1/2キナーゼ、PI‐3‐キナーゼ、c‐Src、R‐Ras、Rac活性化とCdc40依存性パスウェイに導くようです。OxPLによる特定の蛋白質の誘導は、EGR‐1、NFAT,CREB,STAT3およびSREBPを含むいくつかの転写因子により媒介されます。OxPLは慢性血管炎症の単に副産物であるばかりでなく重要なメディエーターであることを、これら最近の知見が示唆しています。
VN Bochkov, phD, Department of Vascular Biology and Thrombosis Research, Vienna University , Vienna, Austria
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
| Oxidized PAPC |
HCB | HC4035-1MG | 1 MG |
¥97,000 |
| Oxidized PAPC |
HCB | HC4036-5MG | 5 MG |
¥256,000 |
商品は「研究用試薬」です。人や動物の医療用・臨床診断用・食品用としては使用しないように、十分ご注意ください。
※ 表示価格について

お問い合わせ
商品に関するご相談・お問い合わせ
![]()
受付時間:平日 (9:00〜17:30)








© COSMO BIO





