このページを印刷する
このページを印刷するDNAのメチル化
多くの真核生物ではゲノム中のCpGジヌクレオチドのシトシンの5位炭素原子にメチル基が付加されます。ゲノム中のCpGジヌクレオチドの60〜90%がメチル化を受けていますが、多くの遺伝子のプロモーター領域にあるCGの豊富な領域(CpGアイランド)ではメチル化を受けていません。
ヒストンH3K9のメチル化が引き金になりDNAメチル化酵素がリクルートされ、DNAのメチル化が誘導されることもわかっています。このように、エピジェネティクスを担ういくつかの機構は、相互に協調し複雑なクロマチンの制御を行っています。
ヒストンの修飾
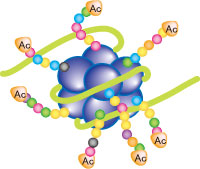
真核生物の染色体DNAはクロマチンとよばれる高次構造をとっています。クロマチンは、ヌクレオソームの繰り返し構造がらせん状につながったものです。ヌクレオソームは、H2A、H2B、H3、H4ヒストンタンパク質が2分子からなるヒストンオクタマーに、146塩基対のDNAが約2回転巻き付いた構造をとっています。
転写誘導の際に、ヒストン修飾によるクロマチン構造変換が重要な働きをすることが知られてきています。
特に転写誘導とよく相関するアセチル化として、ヒストンH3K9とK14のアセチル化が知られています。
ヒストンにはアセチル化とメチル化以外にも様々な修飾を受けることが知られています。リン酸化は細胞分裂時の染色体の凝縮と関係し、ユビキチン化は転写の活性化と、SUMO化は転写の抑制と関係しています。
京都大学ウイルス研究所 生体応答学研究部門 生体防御研究分野 教授 生田 宏一 先生の総説
総説:エピジェネティクス epigenetics - ヒストン修飾とクロマチン構造変換による転写− より抜粋
総説:エピジェネティクス epigenetics - ヒストン修飾とクロマチン構造変換による転写− より抜粋