






















 このページを印刷する
このページを印刷する
記事ID : 8993
G-LISAを用いた心臓研究: 糖尿病性心筋症におけるRho経路に関する研究 CYTOSKELETON NEWS 2012年3月号
G-LISAを用いた心臓研究: 糖尿病性心筋症におけるRho経路に関する研究
Rho/ROCK経路は心血管疾患における心性高血圧の調整に関与しているとされています1。主な効果はアクトミオシンの細胞骨格の張力を調製することにあります2,3,4,5。RhoAの活性化が誘導されることで様々種類の細胞においてアクチン構造の再構成が引き起こされることは十分に立証されています。心組織において、一連の再構成により細胞膜におけるリポプロテインリパーゼ(LPL)の局在の増加が起こり6,7、その結果糖質酸化に代わって脂質代謝(カイロミクロンや超低比重リポタンパクの分解)への依存が高まります8。同様の生理現象が2型糖尿病の高血糖状態においても引き起こされ、糖尿病における細胞の低代謝状態を悪化させます7,8。Rhoタンパクの活性化は細胞骨格の細胞での移動に対する細胞外シグナルの経路の中心となっており、したがってLPLの再局在はRhoタンパクの活性化に依存しています6。LPLの細胞膜への移動を引き起こす経路はアディポネクチン受容体-Rho-ROCK経路とAMPK-p38-MK経路の2種類があると考えられています(Figure 1)。LPL歳局在の生理学的に正常な原因はAMP活性化タンパク質キナーゼ(AMPK)によって検出される低エネルギー状態に対する反応です。このキナーゼはp38 MAPKをリン酸化し、リン酸化されたp38 MAPKは核に移行し、MAPK活性化タンパク質キナーゼ2 (MK2)の翻訳を亢進します。MK2はHSP25リン酸化し、HSP25はアクチン単量体からはずされ、結合のための遊離アクチンの大きなプールを作り出します8。この一連の反応に合わせて、血流中のアディポネクチンがその細胞外受容体に結合しRho/ROCK経路を活性化します。その結果、細胞膜に向かう適切な場所でF-アクチン線維の形成が引き起こされます6。
Figure 1. LPL再局在を制御するシグナル伝達経路
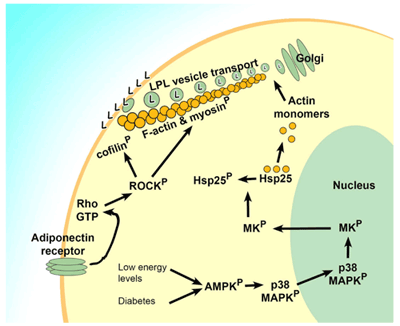
アディポネクチンといった細胞外因子や低エネルギー状態といった細胞内の合図によりAMPKやPho/ROCKを介した経路が誘導され、LPLがゴルジ体から細胞膜に輸送され、細胞骨格の骨組みが形成されます。
CYTOSKELTON社が提供するG-LISAフォーマットは心臓研究において利用可能な少量の心筋細胞や大動脈組織を用いたRhoタンパク活性の測定を可能にしています。このフォーマットには以下のような利点があります4,6,9。1)従来のウェスタンブロット法による解析10に比べ精度が向上します(CV=13%)、2)迅速なプロセッシング、3)少量サンプル(10-50mgタンパク)での解析が可能、4)経 済的である。これらの利点により研究者の方々がこれまでの方法では測定できなかった幅2mm、長さ8mm程度のマウス大動脈といったより小さな組織を用いてRhoタンパクの活性を測定することが可能となります4,6,9。
加えて、ある環境下でGTP-Rhoシグナルを検出限界以下まで下げてしまうリン酸化酵素が高い濃度で混入するために、大動脈ホモジネートを用いたRhoAに関する研究は難しいとされてきました。したがって、大動脈ホモジネートにはGTP-Rhoシグナルが測定に十分な時間安定化するためにリン酸化酵素阻害剤を加える必要がありました。阻害剤には人工基質の混合物や、セリン/スレオニンリン酸化酵素の阻害剤が使用されます。
CYTOSKELTON社では阻害剤として50mM NaF、20mM ピロリン酸ナトリウム、1 mM ミクロシスチン、1mM オルトバナジウム酸を推奨しています。これらの阻害剤を使用しない場合、研究者の方々の中には類似した低い(低下した)GTP-Rhoシグナルを検出することがあります。
最終的には、Rhoタンパク特異的阻害剤および活性剤は神経再生を示すBioaxone社のBA-210(Cethrin®)といった医薬品を使用する傾向にあります。
CYTOSKELTON社ではRho経路研究のための細胞透過性の高いRho阻害剤(Cat. #CT04)と活性剤(Cat. #CN03)を提供しています。
参考文献
1. Hong Z and Yong-Jun L. 2010. RhoA/Rho kinase: a novel therapeutic target in diabetic complications. Chinese Mediacal Journal. 123: 2461-2466.
2. Hirata et al., 1992. Involvement of Rho p21 in the GTP-enhanced calcium ion sensitivity of smooth muscle contraction. J. Biol. Chem., 267, p.8719-8722.
3. Gong et al., 1995. Arachidonic acid and diacylglycerol release associated with inhibition of myosin light chain dephosphorylation... . J. Physiol., 486, p. 113-122.
4. Matsumoto et al., 2010. Enhancement of mesenteric artery contraction to 5-HT depends on Rho kinase and Src kinase pathways in the ob/ob mouse model of type 2 diabetes. Br J Pharmacol., 160(5): 1092–1104.
5. Yang et al., 2012. Mechanism of fibrotic cardiomyopathy in mice expressing truncated Rho-associated coiled-coil protein kinase 1. FASEB J. Jan 25th e-pub ahead of print.
6. Ganguly et al., 2011. Adiponectin increases LPL activity via RhoA/ROCK-mediated actin remodeling in adult rat cardiomyocytes. Endocrinology, 152, p.247-254.
7. Punlinilkunnil and Rodrigues, 2006. Cardiac lipoprotein lipase: Metabolic basis for diabetic heart disease. Cardiovasc. Res., 69, p.329-340.
8. Kim et al., 2007. Acute diabetes moderates trafficking of cardiac lipoprotein lipase through p38 mitogen-activated protein kinase-dependent actin cytoskeleton organization. Diabetes, 57, p.64-76.
9. Seok et al., 2008. Isoflavone Attenuates Vascular Contraction through Inhibition of the RhoA/Rho-Kinase Signaling Pathway. J. Pharmacol. Exp. Ther., 326, p.991–998.
10. Benard and Bokoch, 2002. Assay of Cdc42, Rac, and Rho GTPase activation by affinity methods. Methods Enzymol., 345, p.349-359.
RhoA G-LISA® & ELISAキット
CN01とLPAによるSwiss 3T3細胞Rhoタンパク活性の時間経過による推移
血清不足状態のSwiss 3T3細胞をRho活性剤(Cat. #CN01:青)およびLPA(赤)で処理した。RhoA活性は490nmにおける吸光度により測定した。
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
RhoA G-LISA GTPase Activation Assay (Luminescence format) |
CYT | BK121 | 96 ASSAY |
¥317,000 |
| RhoA G-LISA(R) Activation Assay (absorbance) |
CYT | BK124 | 96 ASSAY |
CYT社 BK124L 1 を参照 |
| Total RhoA ELISA |
CYT | BK150 | 1 KIT [96 assays] |
¥180,000 |
GEF&GAPアッセイ
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
| RhoGEF exchange assay |
CYT | BK100 | 1 KIT [60-300 assays] |
¥240,000 |
| RhoGAP assay |
CYT | BK105 | 1 KIT [80-160 assays] |
¥240,000 |
Rhotekin-RBDビーズ
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
| Rhotekin-RBD protein (binds active Rho proteins), Human |
CYT | RT01-A | 1*500 UG |
¥164,000 |
| Rhotekin-RBD beads (binds active Rho proteins) |
CYT | RT02-A | 2*2 MG |
¥231,000 |
| Rhotekin-RBD beads (binds active Rho proteins) |
CYT | RT02-B | 6*2 MG |
お問い合わせ |
RhoA抗体
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
| Anti RhoA w/positive control, Human (Mouse) |
CYT | ARH03-A | 2*50 UG |
販売終了 |
| Anti RhoA w/positive control, Human (Mouse) |
CYT | ARH03-B | 6*50 UG |
販売終了 |
Rhoアクチベーター&インヒビター
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
| Rho Activator I |
CYT | CN01-A | 5*10 UNIT |
¥76,000 |
| Rho Activator I |
CYT | CN01-B | 20*10 UNIT |
¥268,000 |
| Rho Activator II |
CYT | CN03-A | 3*20 UG |
¥76,000 |
| Rho Activator II |
CYT | CN03-B | 9*20 UG |
¥211,000 |
| Rho/Rac/Cdc42 Activator I |
CYT | CN04-A | 3*20 UG |
¥76,000 |
| Rho/Rac/Cdc42 Activator I |
CYT | CN04-B | 9*20 UG |
¥211,000 |
| Rho Pathway Inhibitor I (Rho Kinase (ROCK) Inhibitor Y-27632) |
CYT | CN06-A | 5*10 UNIT |
販売終了 |
| Rho Pathway Inhibitor I (Rho Kinase (ROCK) Inhibitor Y-27632) |
CYT | CN06-B | 20*10 UNIT |
販売終了 |
| Rho Inhibitor I, Clostridium botulinum , Unlabeled |
CYT | CT04-A | 1*20 UG |
¥73,000 |
| Rho Inhibitor I, Clostridium botulinum , Unlabeled |
CYT | CT04-B | 5*20 UG |
¥237,000 |
Rhoモジュレーター&エフェクタータンパク質
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
| Exoenzyme C3 transferase, Clostridium botulinum , Unlabeled |
CYT | CT03-A | 1*25 UG |
¥73,000 |
| Exoenzyme C3 transferase, Clostridium botulinum , Unlabeled |
CYT | CT03-C | 4*25 UG |
¥264,000 |
| Rho GAP, Human |
CYT | GAP01-A | 1*50 UG |
¥104,000 |
| Rho GAP, Human |
CYT | GAP01-B | 4*50 UG |
¥272,000 |
| Rho GAP, Human |
CYT | GAS01-A | 1*50 UG |
¥104,000 |
| Rho GAP, Human |
CYT | GAS01-B | 4*50 UG |
¥272,000 |
| GDP dissociation inhibitor, Human |
CYT | GDI01-A | 1*25 UG |
¥104,000 |
| Dbs, Human |
CYT | GE01-A | 2*50 UG |
¥104,000 |
Rhoタンパク質
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
| RhoA - constitutively active, Human |
CYT | R6301-A | 1*10 UG |
¥146,000 |
| RhoA - wild type, Human |
CYT | RH01-A | 1*100 UG |
¥146,000 |
| RhoA - wild type, Human |
CYT | RH01-C | 3*100 UG |
¥264,000 |
| RhoA - wild type, Human |
CYT | RH01-XL | 1*1 MG |
お問い合わせ |
| RhoC - wild type, Human |
CYT | RH03-A | 1*100 UG |
¥146,000 |
| RhoA - wild type, Human |
CYT | RHG01-C | 8*25 UG |
¥214,000 |
■ CYTOSKELETON NEWS バックナンバー
- 2020年10月号 紡錘体 - 可視化に向けた新規ツール
- 2020年8月号 細胞膜染色用蛍光プローブ
- 2020年4月号 生細胞におけるF-アクチンプローブ
- 2020年3月号 コロナウイルスと細胞骨格
- 2020年2月号 タウ(Tau)の将来性をMapping
- 2020年1月号 Rho GTPaseによる細胞遊走制御
- 2019年12月号 表現型プロファイリング:アクチンに焦点を当てたがん治療
- 2019年11月号 チューブリンの過剰グルタミル化、ミトコンドリア、神経変性
- 2019年9月号 細胞運動性を制御するために相互作用する膜張力とアクチン細胞骨格
- 2019年8月号 Rac1B、がん、およびRac1
- 2019年7月号 Rhoファミリー GTPases、神経可塑性、およびうつ状態
- 2019年6月号 アクチンメチオニン酸化: 動的制御の次の段階
- 2019年5月号 ミクログリアと神経変性疾患
- 2019年2月号 生細胞画像化に対するCNS疾患や障害
2018年
- 2018年12月号 アクチン細胞骨格とメカノトランスダクション(機械的シグナル伝達)
- 2018年11月号 軸索再生と細胞骨格
- 2018年10月号 ニューロンにおける微小管と極性
- 2018年8月号 Rab GTPase と 神経変性
- 2018年7月号 SUMO レスリング: バランスが全て
- 2018年6月号 なぜ K-Ras は発がん特異性を示すのか?
- 2018年5月号 治療標的としてのユビキチンプロテアソームシステム:チューブリンは関与するか?
- 2018年4月号 RhoファミリーGEFと樹状突起スパインの構造的可塑性
- 2018年3月号 βカテニンとTFC/LEF-1の翻訳後修飾による標準的なWntシグナル制御
- 2018年2月号 がん抑制遺伝子p53の翻訳後修飾による機能の調整
- 2018年1月号 自閉スペクトラム症におけるGEF Trioの役割
2017年
- 2017年12月号 プロフィリン: アクチン結合タンパク質の多機能な役割
- 2017年11月号 ミトコンドリアにおけるアセチル化:新たな考え方と治療への応用の可能性
- 2017年9月号 翻訳後修飾のアセチル化による微小管の安定化
- 2017年8月号 神経軸索におけるアクチンリングを基盤とした周期的膜骨格(PMS)
- 2017年7月号 E3ユビキチンリガーゼMdm2によるがん抑制遺伝子p53の翻訳後制御
- 2017年6月号 多能性幹細胞(PSC)での転写因子による翻訳後制御
- 2017年5月号 Arf6 GEF と癌細胞の浸潤・転移
- 2017年4月号 PTEN(Phosphatase and Tensin Homolog)による翻訳後制御
- 2017年3月号 Tau の翻訳後修飾: アルツハイマー病の治療標的
- 2017年2月号 樹状細胞の移動におけるアクチン結合タンパク質とF-アクチン
2016年
- 2016年11月/12月号 GEF を介した GTPase シグナル伝達の低分子阻害剤
- 2016年9月号 FtsZ タンパク質: 抗菌薬の新規ターゲット
- 2016年7月号 翻訳後修飾(PTM)は心臓病において細胞骨格タンパク質を調節する
- 2016年6月号 モータータンパク質キネシンと神経変性
- 2016年5月号 チロシンリン酸化は Rhoファミリー GTPase 活性を調節する
- 2016年4月号 Rac1と糖尿病: ポジティブな役割とネガティブな役割
- 2016年3月号 SUMO化: 細胞骨格タンパク質の機能を調節するレギュレーター
- 2016年1月/2月号 ビメンチン中間径フィラメント: リン酸化による調節
2015年
- 2015年8月号 タンパク質調節に不可欠な翻訳後修飾
- 2015年7月号 アクチン細胞骨格のライブセルイメージング
- 2015年6月号 有糸分裂に関わるタンパク質のSUMO化: 局在と機能
- 2015年5月号 Ras 癌の治療: 5つの有望なターゲット
- 2015年4月号 Ras 依存性の癌で注目される YAP1
- 2015年3月号 増刊号 統合失調症において遺伝子変異により誘導されるアクチン依存のシナプスの変化
- 2015年3月号 Ral GTPase を調節する翻訳後修飾
- 2015年1月/2月号 RhoA のリン酸化はシグナル伝達を調節する
- 2015年1月号 増刊号 微小管を不安定化する suprafenacine: 新規抗癌剤のリード化合物としての可能性
2014年
- 2014年12月号 増刊号 RhoA は心筋細胞におけるアクチン細胞骨格の再構成とグルコース取り込みを仲介する
- 2014年11月号 増刊号 樹状突起の形態形成: ドーパミンD1受容体 および Rho ファミリー GTPase による制御
- 2014年11月/12月号 GTPase 活性化アッセイ: アイソフォームの検出
- 2014年10月号 アルギニンの正電荷を消失させるシトルリン化
- 2014年9月号 キネシンサブドメインの探索
- 2014年9月号 増刊号 アクチン結合タンパク質コフィリンの S-ニトロシル化: 細胞移動に対する影響
- 2014年8月号 増刊号 原発性硬化性胆管炎における N-Ras 発現および活性
- 2014年8月号 SUMO化: 細胞骨格タンパク質を標的とした翻訳後修飾
- 2014年7月号 Sos/K-Ras 結合を介して Ras シグナル伝達を制御する新しい低分子阻害剤
- 2014年6月号 増刊号 頭頸部扁平上皮癌における microRNA-138 による RhoC のダウンレギュレーション
- 2014年6月号 Rho GTPase と活性酸素種: クロストークとフィードバック
- 2014年5月号 ミオシンのアセチル化はサルコメアの構造と機能を調節する
- 2014年4月号 リジンのアセチル化 - 多様な細胞プロセスの制御因子
- 2014年3月号 インテグリンを介したβ-アクチンの酸化還元制御: PDIの出現
- 2014年1/2月号 ダイニン: 一つのモーターが関わる複数の神経変性疾患
2013年
- 2013年11/12月号 ダイニン:チームとして強力に作用するモータータンパク質
- 2013年10月号 神経変性:Rhes、SUMO化、ハンチントン病
- 2013年9月号 モノユビキチン化:タンパク質調節のダイナミックなタグ
- 2013年8月号 Ras及びRhoのプレニル化による翻訳後修飾:癌創薬における役割
- 2013年7月号 アクチンが引き起こす膜突起による浸潤:コルタクチン
- 2013年6月号 アクチン修飾と細胞骨格
- 2013年5月号 微小管内部の実体
- 2013年4月号 神経変性におけるTauの多面性
- 2013年3月号 蛍光フィブロネクチンタンパク質を用いた特発性肺線維症の創薬
- 2013年1/2月号 樹状突起棘:発生におけるArf6の役割
2012年
- 2012年11/12月号 ミオシンの小分子モジュレーター
- 2012年10月号 Rhoファミリーパスウェイのユビキチン化と制御
- 2012年9月号 神経変性におけるRac1 GTPaseの機能
- 2012年8月号 上皮間葉転換(EMT)とRhoファミリー低分子量G-タンパク質の関与
- 2012年7月号 チューブリンの多重修飾:グルタミル化とグリシル化
- 2012年6月号 細胞接着のフィブロネクチン制御と原線維形成
- 2012年5月号 アクチン酸化サイクルの機能
- 2012年4月号 トラフィッキング:ArfとCdc42/Racの結合
- 2012年3月号 G-LISAを用いた心臓研究: 糖尿病性心筋症におけるRho経路に関する研究
- 2012年1月/2月号 FtsZ: 新たな抗生剤の標的となるチュ−ブリンホモログ
商品は「研究用試薬」です。人や動物の医療用・臨床診断用・食品用としては使用しないように、十分ご注意ください。
※ 表示価格について














