






















 このページを印刷する
このページを印刷する
記事ID : 35069
アクチン細胞骨格とメカノトランスダクション(機械的シグナル伝達) CYTOSKELETON NEWS 2018年12月号
アクチン細胞骨格とメカノトランスダクション(機械的シグナル伝達)
メカノトランスダクション(機械的シグナル伝達)は、機械(物理的など)力に対して特異的細胞応答を誘発する生化学的シグナルへの変換を通じて、細胞が感知、解釈、および応答する複数の段階からなる生物学的プロセスです。本来この応答は、単量体(球状; G-)アクチン、らせん状の重合体(線維状; F-)アクチン、およびアクチン結合タンパク質(ABPs)から構成されるアクチン細胞骨格の再構築を必要とする細胞の突出や退縮を産生するための力産生が関与するため、しばしば機械的です1-3。ABPsは、F-アクチンを葉状仮足、張力線維、糸状仮足、ポドソーム、アクチン星状体、渦、および星型といった多様な構造形態へとダイナミックに組織化します2-4。これらの様々な構造は、細胞の機械的刺激に対する多重応答においてそれぞれ特化した役割を担います。F-アクチンは主に接着点や接着結合への結合を介してこれらのシグナルを伝達し、これにより細胞のアクチン細胞骨格と細胞外マトリックスや他の細胞との接触がそれぞれ調和します5-7(図1)。機械的シグナル伝達におけるアクチン細胞骨格の役割を理解することで、アクチンが関与する細胞構造や機能の力誘導性変化の根底となる従来の基礎生物学の範囲を超えると考えられます。ABPs変異体発現に由来する疾患では、細胞は機械力に対して生理的な応答ができません3, 8-13。本ニュースレターでは、機械的シグナル伝達におけるアクチン細胞骨格の役割を協議します。
アクチン細胞骨格は細胞に与えられた張力に対してメカノセンサー(mechanosensor)として機能します3。疑問なのは、細胞骨格は機械的張力に対してどのように応答するのか、ということです(図1)。様々なアクチンを基にした構造内ではアクチン線維が機械的負荷(線維ごと)に耐えており、この点は電子顕微鏡、蛍光共鳴エネルギー移動(FRET)、原子間力顕微鏡、および光学トラップをはじめとする数々の多様な顕微鏡技術を用いて研究されてきました3, 14。これらの様々なアクチン構造は特定の機械的負荷に付随しており、構造の特定化された細胞機能に対して最適化されています。線維の機械的負荷(張力の増大)によりその立体構造15や、ABPsが線維に結合し影響を及ぼす方法3 が変化します。F-アクチンに関与するタンパク質であるコフィリンの場合、線維の長さが変化するとその結合や機能が影響を受けます。細胞を伸展させる引っ張り力は伸びる方向と平行した線維の長さ応じて増大します。この状況下では、コフィリンの結合親和性が低減し、ミオシンIIの結合親和性は増大します16-18。このようなF-アクチン長や結合パートナーの機械的誘発性変化によりF-アクチンが安定化し、細胞の機械的シグナル伝達過程において根幹をなす張力線維がより簡単に形成されます7, 16-19(図1)。アクチン構造動態における張力誘導性変化は、Arp2/320といったアクチン核形成タンパク質の結合にも影響を及ぼします。アクチンを基にした構造の機械的誘発性変化は、少なくともいくつかの細胞腫において遺伝子発現にも影響を及ぼします。機械的刺激を受ける中でより多くの張力線維が形成されると、転写共役因子YAPが核に転位して活性化されます。YAPはHippoシグナル伝達に不可欠であり、細胞増殖や分化に関与する遺伝子の発現増大を媒介します。したがって、細胞外の機械的な力に対するアクチン細胞骨格の応答により生理学的かつ病態生理学的関連性をもつプロセスをもたらすと考えられます7, 21, 22。
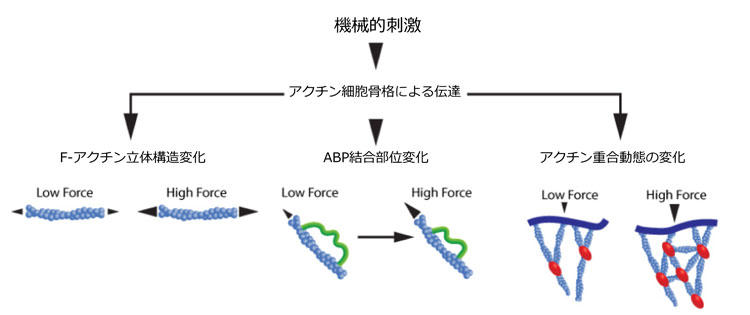
図.1 アクチン細胞骨格は機械的な力を伝達する。
機械的負荷により、1) F-アクチンにおける立体構造変化(左図)、2) ABPsの立体構造変化により隠されていた結合部位があらわになる(中央)、3) ABP介在性アクチン重合動態の変化(右図)、が誘導される。
F-アクチンと同様に、ABPsや他のアクチン関連タンパク質は機械的ストレスに直接応答します(図1)。一般的な応答として立体構造変化が挙げられ、それまで隠されていたタンパク質結合部位があらわになります。接着点関連タンパク質であるタリン、接着結合タンパク質であるα-カテニン、および、ビンキュリン、フィラミン、ミオシン、α-アクチニン4、およびアクチン線維関連タンパク質といったABPsの場合はこれに該当します23-29。
最後に、アクチン重合化とネットワーク集合は、機械力により調節されています(図1)。ABPsにおける力誘導性変化を介して(上記参照)、重合動力学が変化します3。機械力自体も物理的障壁として作用することで重合化に対立できます。重合動力学におけるこのような変化により線維の密度や組織を変更することができるのです3。
まとめ
アクチンは典型的な細胞骨格タンパク質であり、おそらく、細胞の形状、運動性、細胞内輸送、および力産生に変化を生ずる多様な外部刺激に対する細胞応答に最も関連するタンパク質でしょう。アクチンが生理学的かつ病態生理学的プロセスにおいて重要なことは明らかであり、何十年にもわたりそれに焦点を当てた研究がなされてきたのにも関わらず、どのようにしてこれほど多様な高次アクチン構造が細胞内に存在しているのか、また、機械的シグナル伝達においてアクチンに付随する機能は何か、理解することは未だに課題です。これらの疑問に答えるには、線維に対する機械的負荷を適用し測定する方法を組み合わせた高解像度顕微鏡が必要となり、技術的に難易度の高いことです。研究者の方々が機械的シグナル伝達におけるF-アクチンの役割(および、他の細胞プロセス)を明らかにするためのお役に立てるよう、Cytoskeleton, Inc.では精製した標識済み、または未標識アクチンタンパク質、精製ABPs、機能性アクチンアッセイキット、およびF-アクチン生細胞画像化プローブをご提供しています。
参考文献
- Alonso J.L. and Goldmann W.H. 2016. Cellular mechanotransduction. AIMS Biophysics. 3, 50-62.
- Wang N. 2017. Review of cellular mechanotransduction. J. Phys. D. Appl. Phys. 50(23). pii: 233002.
- Harris A.R. et al. 2018. Mechanotransduction by the actin cytoskeleton: Converting mechanical stimuli into biochemicals signals. Annu. Rev. Biophys. 47, 617-631.
- Fritzsche M. et al. 2017. Self-organizing actin patterns shape membrane architecture but not cell mechanics. Nat. Commun. 8, 14347.
- Perez-Moreno M. et al. 2003. Sticky business: orchestrating cellular signals at adherens junctions. Cell. 112, 535-548.
- Geiger B. et al. 2009. Environmental sensing through focal adhesions. Nat. Rev. Mol. Cell Biol. 10, 21-33.
- Ohashi K. et al. 2017. Roles of the cytoskeleton, cell adhesion and rho signaling in mechanosensing and mechanotransduction. J. Biochem. 161, 245-254.
- Jaalouk D.E. and Lammerding J. 2009. Mechanotransduction gone awry. Nat. Rev. Mol. Cell Biol. 10, 63-73.
- Weins A. et al. 2007. Disease-associated mutant α-actinin-4 reveals a mechanism for regulating its F-actin-binding affinity. Proc. Natl. Acad. Sci. USA. 104, 16080-16085.
- Lee S.H. et al. 2008. Crystal structure of the actin-binding domain of α-actinin-4 Lys255Glu mutant implicated in focal segmental glomerulosclerosis. J. Mol. Biol. 376, 317-324.
- Clark A.R. et al. 2009. Skeletal dysplasias due to filamin A mutations result from a gain-of-function mechanism distinct from allelic neurological disorders. Hum. Mol. Genet. 18, 4791-4800.
- Henderson D.M. et al. 2009. Patients with ACTN4 mutations demonstrate distinctive features of glomerular injury. J. Am. Soc. Nephrol. 20, 961-968.
- Henderson D.M. et al. 2010. Disease-causing missense mutations in actin binding domain 1 of dystrophin induce thermodynamic instability and protein aggregation. Proc. Natl. Acad. Sci. USA. 107, 9632-9637.
- Wang Y. and Kanchanawong P. 2016. Three-dimensional super resolution microscopy of F-actin filaments by interferometric photoactivated localization microscopy (iPALM). JoVE. 118, e54774.
- Shimozawa T. and Ishiwata S. 2009. Mechanical distortion of single actin filaments induced by external force: detection by fluorescence imaging. Biophys. J. 96, 1036-1044.
- Uyeda T.Q. et al. 2011. Stretching actin filaments within cells enhances their affinity for the myosin II motor domain. PLoS One. 6, e26200.
- Hayakawa K. et al. 2011. Actin filaments function as a tension sensor by tension-dependent binding of cofilin to the filament. J. Cell Biol. 195, 721-727.
- Hayakawa K. et al. 2014. Single-molecule imaging and kinetic analysis of cooperative cofilin-actin filament interactions. Proc. Natl. Acad. Sci. USA. 111, 9810-9815.
- McGough A. et al. 1997. Cofilin changes the twist of F-actin: implications for actin filament dynamics and cellular function. J. Cell Biol. 138, 771-781.
- Risca V.I. et al. 2012. Actin filament curvature biases branching direction. Proc. Natl. Acad. Sci. USA. 109, 2913-2918.
- Dupont S. et al. 2011. Role of YAP/TAZ in mechanotransduction. Nature. 474, 179-183.
- Halder G. et al. 2012. Transduction of mechanical and cytoskeletal cues by YAP and TAZ. Nat. Rev. Mol. Cell Biol. 13, 591-600.
- Han B. et al. 2004. Conversion of mechanical force into biochemical signaling. J. Biol. Chem. 279, 54793-54801.
- del Rio A. et al. 2009. Stretching single talin rod molecules activates vinculin binding. Science. 323, 638-641.
- Ehrlicher A.J. et al. 2011. Mechanical strain in actin networks regulates FilGAP and integrin binding to filamin A. Nature. 478, 260.
- Luo T. et al. 2013. Molecular mechanisms of cellular mechanosensing. Nat. Mater. 12, 1064-1071.
- Buckley C.D. et al. 2014. The minimal cadherin-catenin complex binds to actin filaments under force. Science. 346, 1254211.
- Huelsmann S. et al. 2016. Evidence for the mechanosensory function of filamin in tissue development. Sci. Rep. 6, 32798.
- Schiffhauer E.S. et al. 2016. Mechanoaccumulative elements of the mammalian actin cytoskeleton. Curr. Biol. 26, 1473-1479.
Actin Products
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
Actin, Bovine, Unlabeled |
CYT | AD99-A | 1*1 MG |
¥44,000 |
| Actin, Bovine, Unlabeled |
CYT | AD99-B | 5*1 MG |
¥181,000 |
| Actin, Chicken |
CYT | AS99-A | 1*1 MG |
¥44,000 |
| Actin, Chicken |
CYT | AS99-B | 5*1 MG |
¥181,000 |
| Actin protein (pre-formed filaments), Rabbit |
CYT | AKF99-A | 1*1 MG |
¥69,000 |
| Actin protein (pre-formed filaments), Rabbit |
CYT | AKF99-B | 5*1 MG |
¥240,000 |
| Actin, Rabbit, Unlabeled |
CYT | AKL95-B | 1*1 MG |
¥35,000 |
| Actin, Rabbit, Unlabeled |
CYT | AKL95-C | 5*1 MG |
¥137,000 |
| Actin, Rabbit, Unlabeled |
CYT | AKL99-A | 4*250 UG |
¥73,000 |
| Actin, Rabbit, Unlabeled |
CYT | AKL99-B | 2*1 MG |
¥87,000 |
| Actin, Rabbit, Unlabeled |
CYT | AKL99-C | 5*1 MG |
¥177,000 |
| Actin, Rabbit, Unlabeled |
CYT | AKL99-D | 10*1 MG |
¥335,000 |
| Actin, Rabbit, Unlabeled |
CYT | AKL99-E | 20*1 MG |
お問い合わせ |
| Actin, Human, Unlabeled |
CYT | APHL99-A | 2*250 UG |
¥79,000 |
| Actin, Human, Unlabeled |
CYT | APHL99-C | 1*1 MG |
¥133,000 |
| Actin, Human, Unlabeled |
CYT | APHL99-E | 5*1 MG |
お問い合わせ |
| Actin, Human, Rhodamine Isothiocyanate |
CYT | APHR-A | 4*10 UG |
¥108,000 |
| Actin, Human, Rhodamine Isothiocyanate |
CYT | APHR-C | 20*10 UG |
¥339,000 |
| Actin, Rabbit, Rhodamine Isothiocyanate |
CYT | AR05-B | 10*20 UG |
¥104,000 |
| Actin, Rabbit, Rhodamine Isothiocyanate |
CYT | AR05-C | 20*20 UG |
¥200,000 |
| Acti-stain 488 phalloidin, Plant |
CYT | PHDG1-A | 1*500 UL [300 slides] |
¥79,000 |
| Acti-stain 555 phalloidin, Mushroom |
CYT | PHDH1-A | 1*500 UL [300 slides] |
¥79,000 |
| Acti-stain 670 phalloidin, Plant |
CYT | PHDN1-A | 1*500 UL [300 slides] |
¥79,000 |
| Rhodamine Phalloidin, Rhodamine Isothiocyanate |
CYT | PHDR1 | 1*500 UL [300 slides] |
¥79,000 |
Actin Biochem Kits
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
| Actin Binding Protein Spin-Down Assay Biochem Kit, Rabbit |
CYT | BK001 | 1 KIT [30-100 assays] |
¥268,000 |
| Actin Binding Protein Spin-Down Assay Biochem Kit, Human |
CYT | BK013 | 1 KIT [30-100 assays] |
¥272,000 |
| Actin Polymerization Biochem Kit (fluorescence format), Rabbit |
CYT | BK003 | 1 KIT [30-100 assays] |
¥304,000 |
| G-Actin/F-actin In Vivo Assay Biochem Kit, Mouse |
CYT | BK037 | 1 KIT [30-100 assays] |
¥264,000 |
■ CYTOSKELETON NEWS バックナンバー
- 2020年10月号 紡錘体 - 可視化に向けた新規ツール
- 2020年8月号 細胞膜染色用蛍光プローブ
- 2020年4月号 生細胞におけるF-アクチンプローブ
- 2020年3月号 コロナウイルスと細胞骨格
- 2020年2月号 タウ(Tau)の将来性をMapping
- 2020年1月号 Rho GTPaseによる細胞遊走制御
- 2019年12月号 表現型プロファイリング:アクチンに焦点を当てたがん治療
- 2019年11月号 チューブリンの過剰グルタミル化、ミトコンドリア、神経変性
- 2019年9月号 細胞運動性を制御するために相互作用する膜張力とアクチン細胞骨格
- 2019年8月号 Rac1B、がん、およびRac1
- 2019年7月号 Rhoファミリー GTPases、神経可塑性、およびうつ状態
- 2019年6月号 アクチンメチオニン酸化: 動的制御の次の段階
- 2019年5月号 ミクログリアと神経変性疾患
- 2019年2月号 生細胞画像化に対するCNS疾患や障害
2018年
- 2018年12月号 アクチン細胞骨格とメカノトランスダクション(機械的シグナル伝達)
- 2018年11月号 軸索再生と細胞骨格
- 2018年10月号 ニューロンにおける微小管と極性
- 2018年8月号 Rab GTPase と 神経変性
- 2018年7月号 SUMO レスリング: バランスが全て
- 2018年6月号 なぜ K-Ras は発がん特異性を示すのか?
- 2018年5月号 治療標的としてのユビキチンプロテアソームシステム:チューブリンは関与するか?
- 2018年4月号 RhoファミリーGEFと樹状突起スパインの構造的可塑性
- 2018年3月号 βカテニンとTFC/LEF-1の翻訳後修飾による標準的なWntシグナル制御
- 2018年2月号 がん抑制遺伝子p53の翻訳後修飾による機能の調整
- 2018年1月号 自閉スペクトラム症におけるGEF Trioの役割
2017年
- 2017年12月号 プロフィリン: アクチン結合タンパク質の多機能な役割
- 2017年11月号 ミトコンドリアにおけるアセチル化:新たな考え方と治療への応用の可能性
- 2017年9月号 翻訳後修飾のアセチル化による微小管の安定化
- 2017年8月号 神経軸索におけるアクチンリングを基盤とした周期的膜骨格(PMS)
- 2017年7月号 E3ユビキチンリガーゼMdm2によるがん抑制遺伝子p53の翻訳後制御
- 2017年6月号 多能性幹細胞(PSC)での転写因子による翻訳後制御
- 2017年5月号 Arf6 GEF と癌細胞の浸潤・転移
- 2017年4月号 PTEN(Phosphatase and Tensin Homolog)による翻訳後制御
- 2017年3月号 Tau の翻訳後修飾: アルツハイマー病の治療標的
- 2017年2月号 樹状細胞の移動におけるアクチン結合タンパク質とF-アクチン
2016年
- 2016年11月/12月号 GEF を介した GTPase シグナル伝達の低分子阻害剤
- 2016年9月号 FtsZ タンパク質: 抗菌薬の新規ターゲット
- 2016年7月号 翻訳後修飾(PTM)は心臓病において細胞骨格タンパク質を調節する
- 2016年6月号 モータータンパク質キネシンと神経変性
- 2016年5月号 チロシンリン酸化は Rhoファミリー GTPase 活性を調節する
- 2016年4月号 Rac1と糖尿病: ポジティブな役割とネガティブな役割
- 2016年3月号 SUMO化: 細胞骨格タンパク質の機能を調節するレギュレーター
- 2016年1月/2月号 ビメンチン中間径フィラメント: リン酸化による調節
2015年
- 2015年8月号 タンパク質調節に不可欠な翻訳後修飾
- 2015年7月号 アクチン細胞骨格のライブセルイメージング
- 2015年6月号 有糸分裂に関わるタンパク質のSUMO化: 局在と機能
- 2015年5月号 Ras 癌の治療: 5つの有望なターゲット
- 2015年4月号 Ras 依存性の癌で注目される YAP1
- 2015年3月号 増刊号 統合失調症において遺伝子変異により誘導されるアクチン依存のシナプスの変化
- 2015年3月号 Ral GTPase を調節する翻訳後修飾
- 2015年1月/2月号 RhoA のリン酸化はシグナル伝達を調節する
- 2015年1月号 増刊号 微小管を不安定化する suprafenacine: 新規抗癌剤のリード化合物としての可能性
2014年
- 2014年12月号 増刊号 RhoA は心筋細胞におけるアクチン細胞骨格の再構成とグルコース取り込みを仲介する
- 2014年11月号 増刊号 樹状突起の形態形成: ドーパミンD1受容体 および Rho ファミリー GTPase による制御
- 2014年11月/12月号 GTPase 活性化アッセイ: アイソフォームの検出
- 2014年10月号 アルギニンの正電荷を消失させるシトルリン化
- 2014年9月号 キネシンサブドメインの探索
- 2014年9月号 増刊号 アクチン結合タンパク質コフィリンの S-ニトロシル化: 細胞移動に対する影響
- 2014年8月号 増刊号 原発性硬化性胆管炎における N-Ras 発現および活性
- 2014年8月号 SUMO化: 細胞骨格タンパク質を標的とした翻訳後修飾
- 2014年7月号 Sos/K-Ras 結合を介して Ras シグナル伝達を制御する新しい低分子阻害剤
- 2014年6月号 増刊号 頭頸部扁平上皮癌における microRNA-138 による RhoC のダウンレギュレーション
- 2014年6月号 Rho GTPase と活性酸素種: クロストークとフィードバック
- 2014年5月号 ミオシンのアセチル化はサルコメアの構造と機能を調節する
- 2014年4月号 リジンのアセチル化 - 多様な細胞プロセスの制御因子
- 2014年3月号 インテグリンを介したβ-アクチンの酸化還元制御: PDIの出現
- 2014年1/2月号 ダイニン: 一つのモーターが関わる複数の神経変性疾患
2013年
- 2013年11/12月号 ダイニン:チームとして強力に作用するモータータンパク質
- 2013年10月号 神経変性:Rhes、SUMO化、ハンチントン病
- 2013年9月号 モノユビキチン化:タンパク質調節のダイナミックなタグ
- 2013年8月号 Ras及びRhoのプレニル化による翻訳後修飾:癌創薬における役割
- 2013年7月号 アクチンが引き起こす膜突起による浸潤:コルタクチン
- 2013年6月号 アクチン修飾と細胞骨格
- 2013年5月号 微小管内部の実体
- 2013年4月号 神経変性におけるTauの多面性
- 2013年3月号 蛍光フィブロネクチンタンパク質を用いた特発性肺線維症の創薬
- 2013年1/2月号 樹状突起棘:発生におけるArf6の役割
2012年
- 2012年11/12月号 ミオシンの小分子モジュレーター
- 2012年10月号 Rhoファミリーパスウェイのユビキチン化と制御
- 2012年9月号 神経変性におけるRac1 GTPaseの機能
- 2012年8月号 上皮間葉転換(EMT)とRhoファミリー低分子量G-タンパク質の関与
- 2012年7月号 チューブリンの多重修飾:グルタミル化とグリシル化
- 2012年6月号 細胞接着のフィブロネクチン制御と原線維形成
- 2012年5月号 アクチン酸化サイクルの機能
- 2012年4月号 トラフィッキング:ArfとCdc42/Racの結合
- 2012年3月号 G-LISAを用いた心臓研究: 糖尿病性心筋症におけるRho経路に関する研究
- 2012年1月/2月号 FtsZ: 新たな抗生剤の標的となるチュ−ブリンホモログ
商品は「研究用試薬」です。人や動物の医療用・臨床診断用・食品用としては使用しないように、十分ご注意ください。
※ 表示価格について














