






















 このページを印刷する
このページを印刷する
記事ID : 10510
樹状突起棘:発生におけるArf6の役割 CYTOSKELETON NEWS 2013年1月/2月号
樹状突起棘:発生におけるArf6の役割
樹状突起は、形成、伸長、分岐を経て発生します。特に興奮性シナプスを持つ神経の発生は、樹状突起の糸状仮足(樹状突起棘の前駆体)のようなアクチンの突出の過程を経ます(図1)。糸状仮足は、樹状突起棘の前駆体である一方、全ての糸状仮足が樹状突起棘に分化するわけではありません。構造的に、樹状突起棘は主にF-アクチンで構成され、大部分の興奮性シナプスの神経伝達の場となります1-4。樹状突起棘の構造及び機能のダイナミックな変化は、正常な学習と記憶の過程だけでなく、神経変性や神経学的障害に関係する疾病(例:アルツハイマー病(AD)、精神遅滞)の過程でも重要な役割を果たします1,5,6。従って、樹状突起及び棘中のアクチン動力を調節する経路は、基礎・臨床両方の神経科学者から大きな関心を集めています。樹状突起棘の形態、可動性、安定性(及びそれらに関する機能)を調節するために必要な2つのプロセスは、細胞骨格アクチンリモデリング3,4と膜輸送7です。
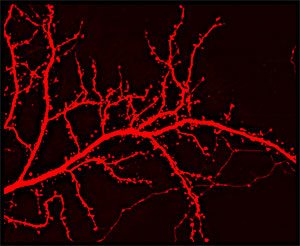
図1 DsRedをトランスフェクトした海馬ニューロンの樹状突起と棘
低分子量Gタンパク質のRasスーパーファミリーの一部であるADPリボシル化因子(Arf)GTPaseは、アクチンの再編成や輸送の機能を持つことから、神経細胞の樹状突起及び棘の発生に重要な役割を果たしていることが、最近明らかになりました。他のGTPaseと同様に、Arfはグアニン交換因子(GEFs)によって媒介されるGTP結合状態と、グアニン活性化タンパク質(GAPs)によって媒介されるGDP結合状態を循環しています。Arfの6つの既知のアイソフォーム8,9のうち、Arf6は他のArfとは異なっており、独特の機能と細胞内局在性9を持ちます。Arf6は、神経細胞を含むほとんどの細胞で、輸送とアクチンの再編成という2つの機能を持っています7。GTP結合Arf6は細胞膜に局在するのに対し、GDP結合Arf6はエンドソームの区画に局在します9。
Arfと樹状突起の分岐
海馬の神経細胞を用いた初期の樹状細胞の発生の研究で、Arf6は樹状細胞の分岐を、少なくとも2つの経路によってネガティブに調節している10-12ことが報告されました。1つはArfのエフェクターを介する直接的な経路、もう一方はRac1を介する間接的な経路です10(図2)。Arf6 GEFのドミナントネガティブ(DN)変異の過剰発現は、樹状突起の分岐を著しく増加させます12。これはおそらくαアクチンが関わる機構が影響していると思われます13。同様に、Arf GAPのsiRNAノックダウン又はGAPのDNは、樹状突起の分岐を減少させ、野生型のGAPの過剰発現は分岐を増加させます14。
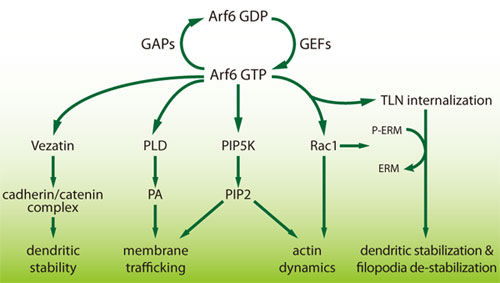
図2 Arf6シグナル伝達経路
PLD:ホスホリパーゼ D、PA:ホスファチジン酸、PIP2:ホスファチジルイノシトールビスリン酸、PIP5K:ホスファチジルイノシトール-4-リン酸塩5-キナーゼ、TLN:テレンセファリン
Arfと樹状突起棘形成
樹状突起の発生の最終段階は、シナプスの形成です。興奮性のシナプスの場合は糸状仮足の形成を経て樹状突起棘へと発生します。野生型の過剰発現又は恒常活性型(CA)のArf6による研究で、Arf6が樹状突起の糸状仮足の形成をネガティブに調節していることが明らかになりました11。同様の実験で、培養海馬神経細胞中の樹状突起棘の密度が減少する一方で、Arf6のDNの過剰発現は反対の効果をもたらしました15。同様に、海馬切片の培養においても、Arf GAPのDNの過剰発現が樹状突起棘の密度を減少させました14。反対に、Choiらは、fast-cycling Arf変異体(CA)の過剰発現、もしくは野生型Arf GEFでは、樹状突起棘の数が増加し、Arf変異体では成熟神経細胞中の糸状仮足の数が減少すると報告しました16。さらに、Arf6またはArf GEFのsiRNAノックダウンにより、樹状突起棘の数が減少する一方で、糸状仮足の数が増加することがわかりました。生細胞の画像により、この変化は、糸状仮足から樹状突起棘への転換が減少しているためであることが確認されました。また、Rac1の活性化は糸状仮足から樹状突起棘への発達に関与していることも報告されています16。Raemaekersら17は、同じfast-cycling Arf変異体を過剰発現させた海馬神経細胞で、糸状仮足の増加に伴い樹状突起棘の数が減少することを確認しました。これらの研究は、培養神経細胞に対してArf GEF及びArf GAPを外因的に発現させています。矛盾したデータの要因として、異なる変異体が使われたこと、GEFsとGAPsが複数のArfsを管理していることから、対立する機能的な影響を与えた可能性が考えられます。加えて、外因的に発現されたタンパク質の内因的な結合相手もしくは野生型の対象物との相互作用が、過剰発現されたこれらのタンパク質の機能的な出力を変化させた可能性もあります。また、トランスフェクトされたタンパク質の発現レベルは異なっています。これらの要因が、神経細胞におけるArf GTPaseによる樹状突起と棘の形成の調節に関する矛盾したデータを解明する鍵となるかもしれません。
樹状突起と棘の形成におけるこれらの変化を媒介するArfエフェクターの下流因子はまだ同定されていません(図2)。候補として、ホスファチジルイノシトール-4-リン酸塩5キナーゼ、ホスホリパーゼD、vezatin、Rac1 GTPaseが挙げられます7,18。もう1つの候補であるテレンセファリン(TLN;細胞接着分子5としても知られる)は、Arf6によって内部移行及び輸送を媒介され、樹状突起棘の発達を阻害します17。Arf6が媒介するTLNのエンドサイトーシスは、最終的に樹状突起棘の安定性を増強する経路を惹起します。これは少なくとも部分的にRac1が媒介するERMアクチン結合タンパク質の脱リン酸化によって調節されており、細胞膜からTLNを分離させます17。
Arf6のほかにArf4も樹状突起棘の発達をポジティブに調節します19。Arf4のノックダウン(Arf+/- mice)は培養海馬神経細胞中の樹状突起棘の濃度を減少させる一方、野生型又はCA Arf4の過剰発現は判定の効果を示します。臨床的に、Arf4の過剰発現は、マウスADモデルの海馬神経細胞で、アポリポタンパク質E4の発現により誘導される樹状突起棘の損失を防ぎます19。
これらの進歩にも関わらず、Arfsが樹状突起の発達を調節する方法の正確な解明だけでなく、各種Arf4と6のGEFとGAPへの特異性と重複性についても、多くの疑問はまだ残ったままです。これらの疑問を解決するのを支援するため、サイトスケルトン社は、複数のArf活性化アッセイキットとArf関連試薬をご提供致します。
活性化アッセイキット
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 | 在庫 |
|---|---|---|---|---|---|
Arf1 G-LISA Activation Assay Kit |
CYT | BK132 | 96 ASSAY |
販売終了 | 販売終了 |
| Arf1 Pull-down Activation Assay Biochem Kit (bead pull-down format) |
CYT | BK032-S | 20 ASSAY |
¥170,000 | なし |
| Arf6 G-LISA Activation Assay Kit |
CYT | BK133 | 96 ASSAY |
販売終了 | 販売終了 |
| Arf6 Pull-down Activation Assay Biochem Kit (bead pull-down format) |
CYT | BK033-S | 20 ASSAY |
¥170,000 | なし |
| Rac1 -2 -3 G-LISA GTPase Activation Assay (Colorimetric format) |
CYT | BK125 | 96 ASSAY |
¥317,000 | なし |
| Rac1 G-LISA(R) Activation Assay Kit |
CYT | BK128 | 1 KIT [96 assays] |
CYT社 BK128L 1 を参照 |
なし |
| Rac1 Pulldown Activation Assay Kit |
CYT | BK035 | 1 KIT [50 assays] |
CYT社 BK035L 1 を参照 |
なし |
| Rac1 Pull-down Activation Assay Biochem Kit (bead pull-down format) |
CYT | BK035-S | 1 KIT [20 assays] |
¥132,000 | なし |
タンパク質
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 | 在庫 |
|---|---|---|---|---|---|
| Rac1 - constitutively active, Human |
CYT | R6101-A | 1*10 UG |
¥146,000 | なし |
| Rac1 His Protein: constitutively active |
CYT | R6101-C | 4*10 UG |
販売終了 | 販売終了 |
| Rac1 - dominant negative, Human |
CYT | R17G01-A | 1*25 UG |
¥146,000 | なし |
| Rac1 GST Protein: dominant negative, Human |
CYT | R17G01-C | 4*25 UG |
販売終了 | 販売終了 |
| Rac1 - wild type, Human |
CYT | RCG01-C | 8*25 UG |
¥231,000 | なし |
| Rac1 - wild type, Human |
CYT | RC01-A | 1*100 UG |
¥146,000 | なし |
| Rac1 - wild type, Human |
CYT | RC01-C | 3*100 UG |
¥264,000 | なし |
| Rac1 - wild type, Human |
CYT | RC01-XL | 1*1 MG |
お問い合わせ | なし |
| Rac2 - wild type, Human |
CYT | RC02-A | 1*100 UG |
¥146,000 | なし |
| Rac2, His tagged |
CYT | RC02-B | 3*100 UG |
販売終了 | 販売終了 |
低分子Gタンパク質抗体
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 | 在庫 |
|---|---|---|---|---|---|
| Anti Rac 1 w/positive control (Mouse) Unlabeled |
CYT | ARC03-A | 2*50 UG |
販売終了 | 販売終了 |
| Anti Rac 1 w/positive control (Mouse) Unlabeled |
CYT | ARC03-B | 6*50 UG |
販売終了 | 販売終了 |
ファロイジン
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 | 在庫 |
|---|---|---|---|---|---|
| Acti-stain 488 phalloidin, Plant |
CYT | PHDG1-A | 1*500 UL [300 slides] |
¥79,000 | あり |
| Rhodamine Phalloidin, Rhodamine Isothiocyanate |
CYT | PHDR1 | 1*500 UL [300 slides] |
¥79,000 | あり |
| Acti-stain 555 phalloidin, Mushroom |
CYT | PHDH1-A | 1*500 UL [300 slides] |
¥79,000 | あり |
| Acti-stain 670 phalloidin, Plant |
CYT | PHDN1-A | 1*500 UL [300 slides] |
¥79,000 | あり |
■ CYTOSKELETON NEWS バックナンバー
- 2020年10月号 紡錘体 - 可視化に向けた新規ツール
- 2020年8月号 細胞膜染色用蛍光プローブ
- 2020年4月号 生細胞におけるF-アクチンプローブ
- 2020年3月号 コロナウイルスと細胞骨格
- 2020年2月号 タウ(Tau)の将来性をMapping
- 2020年1月号 Rho GTPaseによる細胞遊走制御
- 2019年12月号 表現型プロファイリング:アクチンに焦点を当てたがん治療
- 2019年11月号 チューブリンの過剰グルタミル化、ミトコンドリア、神経変性
- 2019年9月号 細胞運動性を制御するために相互作用する膜張力とアクチン細胞骨格
- 2019年8月号 Rac1B、がん、およびRac1
- 2019年7月号 Rhoファミリー GTPases、神経可塑性、およびうつ状態
- 2019年6月号 アクチンメチオニン酸化: 動的制御の次の段階
- 2019年5月号 ミクログリアと神経変性疾患
- 2019年2月号 生細胞画像化に対するCNS疾患や障害
2018年
- 2018年12月号 アクチン細胞骨格とメカノトランスダクション(機械的シグナル伝達)
- 2018年11月号 軸索再生と細胞骨格
- 2018年10月号 ニューロンにおける微小管と極性
- 2018年8月号 Rab GTPase と 神経変性
- 2018年7月号 SUMO レスリング: バランスが全て
- 2018年6月号 なぜ K-Ras は発がん特異性を示すのか?
- 2018年5月号 治療標的としてのユビキチンプロテアソームシステム:チューブリンは関与するか?
- 2018年4月号 RhoファミリーGEFと樹状突起スパインの構造的可塑性
- 2018年3月号 βカテニンとTFC/LEF-1の翻訳後修飾による標準的なWntシグナル制御
- 2018年2月号 がん抑制遺伝子p53の翻訳後修飾による機能の調整
- 2018年1月号 自閉スペクトラム症におけるGEF Trioの役割
2017年
- 2017年12月号 プロフィリン: アクチン結合タンパク質の多機能な役割
- 2017年11月号 ミトコンドリアにおけるアセチル化:新たな考え方と治療への応用の可能性
- 2017年9月号 翻訳後修飾のアセチル化による微小管の安定化
- 2017年8月号 神経軸索におけるアクチンリングを基盤とした周期的膜骨格(PMS)
- 2017年7月号 E3ユビキチンリガーゼMdm2によるがん抑制遺伝子p53の翻訳後制御
- 2017年6月号 多能性幹細胞(PSC)での転写因子による翻訳後制御
- 2017年5月号 Arf6 GEF と癌細胞の浸潤・転移
- 2017年4月号 PTEN(Phosphatase and Tensin Homolog)による翻訳後制御
- 2017年3月号 Tau の翻訳後修飾: アルツハイマー病の治療標的
- 2017年2月号 樹状細胞の移動におけるアクチン結合タンパク質とF-アクチン
2016年
- 2016年11月/12月号 GEF を介した GTPase シグナル伝達の低分子阻害剤
- 2016年9月号 FtsZ タンパク質: 抗菌薬の新規ターゲット
- 2016年7月号 翻訳後修飾(PTM)は心臓病において細胞骨格タンパク質を調節する
- 2016年6月号 モータータンパク質キネシンと神経変性
- 2016年5月号 チロシンリン酸化は Rhoファミリー GTPase 活性を調節する
- 2016年4月号 Rac1と糖尿病: ポジティブな役割とネガティブな役割
- 2016年3月号 SUMO化: 細胞骨格タンパク質の機能を調節するレギュレーター
- 2016年1月/2月号 ビメンチン中間径フィラメント: リン酸化による調節
2015年
- 2015年8月号 タンパク質調節に不可欠な翻訳後修飾
- 2015年7月号 アクチン細胞骨格のライブセルイメージング
- 2015年6月号 有糸分裂に関わるタンパク質のSUMO化: 局在と機能
- 2015年5月号 Ras 癌の治療: 5つの有望なターゲット
- 2015年4月号 Ras 依存性の癌で注目される YAP1
- 2015年3月号 増刊号 統合失調症において遺伝子変異により誘導されるアクチン依存のシナプスの変化
- 2015年3月号 Ral GTPase を調節する翻訳後修飾
- 2015年1月/2月号 RhoA のリン酸化はシグナル伝達を調節する
- 2015年1月号 増刊号 微小管を不安定化する suprafenacine: 新規抗癌剤のリード化合物としての可能性
2014年
- 2014年12月号 増刊号 RhoA は心筋細胞におけるアクチン細胞骨格の再構成とグルコース取り込みを仲介する
- 2014年11月号 増刊号 樹状突起の形態形成: ドーパミンD1受容体 および Rho ファミリー GTPase による制御
- 2014年11月/12月号 GTPase 活性化アッセイ: アイソフォームの検出
- 2014年10月号 アルギニンの正電荷を消失させるシトルリン化
- 2014年9月号 キネシンサブドメインの探索
- 2014年9月号 増刊号 アクチン結合タンパク質コフィリンの S-ニトロシル化: 細胞移動に対する影響
- 2014年8月号 増刊号 原発性硬化性胆管炎における N-Ras 発現および活性
- 2014年8月号 SUMO化: 細胞骨格タンパク質を標的とした翻訳後修飾
- 2014年7月号 Sos/K-Ras 結合を介して Ras シグナル伝達を制御する新しい低分子阻害剤
- 2014年6月号 増刊号 頭頸部扁平上皮癌における microRNA-138 による RhoC のダウンレギュレーション
- 2014年6月号 Rho GTPase と活性酸素種: クロストークとフィードバック
- 2014年5月号 ミオシンのアセチル化はサルコメアの構造と機能を調節する
- 2014年4月号 リジンのアセチル化 - 多様な細胞プロセスの制御因子
- 2014年3月号 インテグリンを介したβ-アクチンの酸化還元制御: PDIの出現
- 2014年1/2月号 ダイニン: 一つのモーターが関わる複数の神経変性疾患
2013年
- 2013年11/12月号 ダイニン:チームとして強力に作用するモータータンパク質
- 2013年10月号 神経変性:Rhes、SUMO化、ハンチントン病
- 2013年9月号 モノユビキチン化:タンパク質調節のダイナミックなタグ
- 2013年8月号 Ras及びRhoのプレニル化による翻訳後修飾:癌創薬における役割
- 2013年7月号 アクチンが引き起こす膜突起による浸潤:コルタクチン
- 2013年6月号 アクチン修飾と細胞骨格
- 2013年5月号 微小管内部の実体
- 2013年4月号 神経変性におけるTauの多面性
- 2013年3月号 蛍光フィブロネクチンタンパク質を用いた特発性肺線維症の創薬
- 2013年1/2月号 樹状突起棘:発生におけるArf6の役割
2012年
- 2012年11/12月号 ミオシンの小分子モジュレーター
- 2012年10月号 Rhoファミリーパスウェイのユビキチン化と制御
- 2012年9月号 神経変性におけるRac1 GTPaseの機能
- 2012年8月号 上皮間葉転換(EMT)とRhoファミリー低分子量G-タンパク質の関与
- 2012年7月号 チューブリンの多重修飾:グルタミル化とグリシル化
- 2012年6月号 細胞接着のフィブロネクチン制御と原線維形成
- 2012年5月号 アクチン酸化サイクルの機能
- 2012年4月号 トラフィッキング:ArfとCdc42/Racの結合
- 2012年3月号 G-LISAを用いた心臓研究: 糖尿病性心筋症におけるRho経路に関する研究
- 2012年1月/2月号 FtsZ: 新たな抗生剤の標的となるチュ−ブリンホモログ
商品は「研究用試薬」です。人や動物の医療用・臨床診断用・食品用としては使用しないように、十分ご注意ください。
※ 表示価格について














