






















 このページを印刷する
このページを印刷する
記事ID : 12847
Rho GTPase と活性酸素種: クロストークとフィードバック CYTOSKELETON NEWS 2014年6月号
Rho GTPase と活性酸素種: クロストークとフィードバック
活性酸素種(ROS)や活性窒素種(RNS)のような酸化・還元剤は、インテグリンシグナル伝達、細胞外マトリックスの接着、炎症などの様々なシグナル伝達経路における重要な制御因子です1-3。Rho GTPase もまた、細胞増殖、運動、接着などの多くの細胞プロセスにおける重要な制御因子です4。酸化・還元剤と Rho GTPase は、多種の制御機構に作用しており、ROS/RNS - Rho GTPase 間のクロストークは、多くの生理機能や5,6、急性肺損傷、癌などの発病、進行過程において7,8、極めて重要な役割を果たしていると考えられます。急速に発展する本分野において、広く知られる知見および最近の新しい報告を以下にまとめます(図1)。
Rho GTPase による ROS の制御
細胞内の ROS/RNS 生成は、主に、NADPH オキシダーゼ(NOX)、一酸化窒素合成酵素(NOS)、スーパーオキシドジスムターゼ(SOD)などの、主要な酸化還元酵素に由来します5,9,10(図1)。Rho GTPase(特に Rac1と Rac2)は、活性化されたNOX 複合体の構成要素であり、続いて起こる酸素分子(O2)からの ROS(スーパーオキシドアニオン、O2.-)生成に必要となります11。細胞外シグナルによって活性化されると、NOX 複合体は細胞膜で会合し、Rac(GTP結合型、おそらくGDIが結合)の関与によりROS が生成されます12,13(図1)。Rac が、アダプターとして作用するのか、O2.- 生成の電子伝達を仲介するのかは、未だ明らかになっていません12。また、Rac(特に Rac1)は、一酸化窒素(NO)の生成を制御する酵素である NOS と、直接相互作用することが示されています13。NOS は、ヌクレオチドフリー型の GTPase に優先的に結合することが報告されており、Rac が仲介する NO 産生に、グアニンヌクレオチド交換因子(GEF)、及び GEF を介した細胞内区画化が関与していることが示唆されます13。さらに、Rac は、O2.- の過酸化水素(H2O2)への変換を触媒する酵素である SOD と、酸化還元によって直接相互作用することが示されています14。還元条件下では、細胞内と同様に、Rac-GTP が SOD に結合することで Rac の活性が維持され、NOX による O2.- の産生が促進されます。酸化条件下では、局所的な O2.- / H2O2 生成と同様に、まだ同定されていない GTPase 活性化タンパク質(GAP)によって、SOD が Rac から解離し、不活性型である GDP 結合型に変換されると考えられています14。これらの報告から、Rho GTPase は、多数の活性酸素中間体(RIs)を生成し、多様なタンパク質修飾をもたらす可能性があることが明確に示されます。生成される ROS/RNS、及び誘発される細胞応答の種類は、与えられた刺激、細胞内の局所的な酸化還元電位、細胞の位置などによると考えられます。
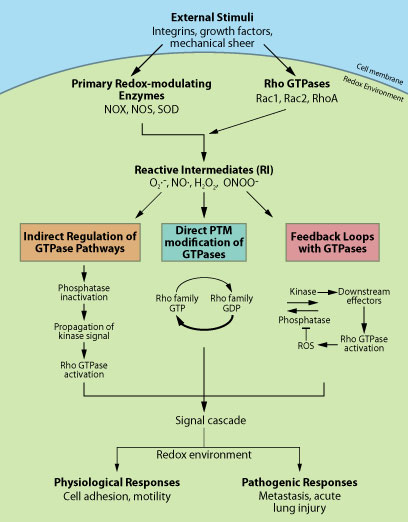
図1 シグナル伝達における Rho GTPase - ROS 間のクロストーク
細胞内において想定される Rho GTPase − ROS/RNS 間のクロストーク概略図。間接的(オレンジ色)、直接的(青色)、フィードバック(ピンク)に分類し、その下に、最も一般的なメカニズムを示した。各経路の詳細については、参考文献および本文中の例を参照。
ROS による直接的/間接的な Rho GTPase の制御
近年、 酸化還元を介した翻訳後修飾(PTMs)によって、ROS/RNS が直接 Rho GTPase を制御する可能性がある、という興味深い報告がなされています。例えば、Heo と Campbell15 は、Rho GTPase(RhoA、Rac、Cdc42 など)の P-ループにある保存配列(GXXXXGK(S/T))の端に位置する、保存されたレドックス感受性モチーフ Cys18 (Rac のナンバリング)が、in vitro で酸化還元を介して可逆的に酸化されることを示しました。この酸化によって、ヌクレオチドの置換が起こり、還元条件下では、GTPヌクレオチド交換や GTPase の活性化が起こります15。このメカニズムは、in vivo においても証明されており、H2O2を介した RhoA の活性化とストレスファイバーの形成が、Cys16 と Cys20 (Rho のナンバリング)の変異によって阻害されることがわかっています16。重要なことに、これらのシステイン変異は、GEFs によって活性化されたRhoA の機能は阻害しません16。筆者らは、GTPase の活性化は、酵素(GEFs/GAPs)及び酸化還元を介した交換の、2つの並行するメカニズムを介して起こることを示唆しています(図1)。また、筆者らは、使用されるメカニズムの決定に、細胞の酸化還元電位が影響すると考えています16。酸化還元を介した交換メカニズムの根拠として、急性肺損傷により増強されたNO産生を介して、RhoA のスイッチ1領域にある Tyr34 がニトロ化されるという報告があります7。筆者らは、このPTMによって、 RhoA が GEF 様のメカニズムで活性化されることを示しました。さらに、酸化還元を介した PTM は、Rho GTPase を不活性化する場合があることも示唆しています17。
直接的な制御に加えて、ホスファターゼが関与する Rho GTPase の間接的なレドックス依存的制御が報告されています。例えば、Rac によって活性化される、 ROS 産生が RhoA の不活性化を引き起こすメカニズムには、酸化還元を介した低分子量タンパク質チロシンホスファターゼ(LMW-PTP)の不活性化、p190 RhoGAP 活性の上昇、RhoA の非活性化が含まれます18。興味深いことに、ROS によるホスファターゼの不活性化は、多様な細胞経路において ROS を制御するための共通のメカニズムであると考えられます19(図1)。
ROS と Rho GTPase 間のフィードバックループ
近年、白血球の走化性の制御に関して20、Rho GTPase が酸化還元のフィードバックループに関与するという仮説が示されています5(図1)。Rac が介する ROS 産生は、酸化還元によるホスファターゼの不活性化を引き起こします。そのことによって、Rac の GEF による活性化を仲介するキナーゼが活性化されます20(図1)。Rac を介する ROS 産生の SOD による制御に関しても、フィードバックのメカニズムが提案されています14。
まとめ
Rho GTPase と酸化・還元剤との関係は、非常に複雑であり、GAP/GEF 制御による交換プロセスと、酸化還元が介する交換プロセスとの関係についての研究は、今後の興味深い分野になると考えられます。2つのメカニズムは並行して、あるいは相乗的に働いていると考えられ、有望な新薬のターゲットとなる可能性があります。
参考文献
1 Yan B. & Smith J.W. 2000. A redox site involved in integrin activation. J. Biol. Chem. 275, 39964-39972.
2 Chiarugi P. et al. 2003. Reactive oxygen species as essential mediators of cell adhesion: the oxidative inhibition of a FAK tyrosine phosphatase is required for cell adhesion. J. Cell Biol. 161, 933-944.
3 Leto T.L. & Geiszt M. 2006. Role of Nox family NADPH oxidases in host defense. Antioxid. Redox Signal. 8, 1549-1561.
4 Jaffe A.B. & Hall A. 2005. Rho GTPases: biochemistry and biology. Annu. Rev. Cell Dev. Biol. 21, 247-269.
5 Mitchell L. et al. 2013. Redox regulation of Ras and Rho GTPases: mechanism and function. Antioxid. Redox Signal.18, 250-258.
6 Ferro E. et al. 2014. Ras GTPases are both regulators and effectors of redox agents. Ras Signaling: Methods Mol. Biol. 1120, 55-74.
7 Rafikov R. et al. 2014. Lipopolysaccharide-induced lung injury involves the nitration-mediated activation of RhoA. J. Biol. Chem. 289, 4710-4722.
8 Myant K.B. et al. 2013. ROS production and NF-kappaB activation triggered by Rac1 facilitate WNT-driven intestinal stem cell proliferation and colorectal cancer initiation. Cell Stem Cell. 12, 761-773.
9 Ueyama T. et al. 2006. Involvement of Rac1 in activation of multicomponent Nox-1 and Nox-3 based NADPH oxidases. Mol. Cell Biol. 26, 2160-2174.
10 Werner E. 2004. GTPases and reactive oxygen species: switches for killing and signaling. J. Cell Sci. 117, 143-153.
11 Hordijk P.L. 2006. Regulation of NADPH oxidases: the role of Rac proteins. Circ. Res. 98, 453-462.
12 Bokoch G.M. 1995. Regulation of the phagocyte respiratory burst by small GTP-binding proteins. Trends Cell Biol.5, 109-113.
13 Di-Poi N. et al. 2001. Mechanism of NADPH oxidase activation by Rac/Rho-GDI complex. Biochemistry. 40, 10014-10022.
14 Harraz M.M. et al. 2008. SOD1 mutations disrupt redox-sensitive Rac regulation of NADPH oxidase in a familial ALS model. J. Clin. Invest. 118, 659-670.
15 Heo J. and Campbell S.L. 2005. Mechanism of redox-mediated guanine nucleotide exchange on redox-active Rho GTPases. J. Biol. Chem. 280, 31003-31010.
16 Aghajanian A. et al. 2009. Direct activation of RhoA by reactive oxygen species requires a redox-sensitive motif.PLoS ONE. 4, e8045.
17 Zuckerbraun B.S. et al. 2007. Nitric oxide-induced inhibition of smooth muscle cell proliferation involves S-nitrosation and inactivation of RhoA. Am. J. Physiol. Cell Physiol. 292, C824-C831.
18 Nimnual A.S. et al. 2003. Redox-dependent downregulation of Rho by Rac. Nat. Cell Biol. 5, 236-241.
19 Meng T.C. et al. 2002. Reversible oxidation and inactivation of protein tyrosine phosphatases in vivo. Mol. Cell. 9, 387-399.
20 Kuiper J.W. et al. 2011. Rac regulates PtdInsP3 signaling and the chemotactic compass through a redox-mediated feedback loop. Blood. 118, 6164-6171.
G-Switch™ アクチベーター/インヒビター
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
Rho Inhibitor I, Clostridium botulinum , Unlabeled |
CYT | CT04-A | 1*20 UG |
¥73,000 |
| Rho Inhibitor I, Clostridium botulinum , Unlabeled |
CYT | CT04-B | 5*20 UG |
¥237,000 |
| Rho Activator II |
CYT | CN03-A | 3*20 UG |
¥76,000 |
| Rho Activator II |
CYT | CN03-B | 9*20 UG |
¥211,000 |
| Rho/Rac/Cdc42 Activator I |
CYT | CN04-A | 3*20 UG |
¥76,000 |
| Rho/Rac/Cdc42 Activator I |
CYT | CN04-B | 9*20 UG |
¥211,000 |
| Rho Pathway Inhibitor I (Rho Kinase (ROCK) Inhibitor Y-27632) |
CYT | CN06-A | 5*10 UNIT |
販売終了 |
| Rho Pathway Inhibitor I (Rho Kinase (ROCK) Inhibitor Y-27632) |
CYT | CN06-B | 20*10 UNIT |
販売終了 |
低分子量Gタンパク質活性化アッセイキット
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
| RhoA Pulldown Activation Assay Kit |
CYT | BK036 | 1 KIT [80 assays] |
CYT社 BK036L 1 を参照 |
| RhoA Pull-down Activation Assay Biochem Kit (bead pull-down format) |
CYT | BK036-S | 1 KIT [20 assays] |
¥132,000 |
| Rac1 Pulldown Activation Assay Kit |
CYT | BK035 | 1 KIT [50 assays] |
CYT社 BK035L 1 を参照 |
| Rac1 Pull-down Activation Assay Biochem Kit (bead pull-down format) |
CYT | BK035-S | 1 KIT [20 assays] |
¥132,000 |
| Cdc42 Pulldown Activation Assay Kit |
CYT | BK034 | 1 KIT [50 assays] |
CYT社 BK034L 1 を参照 |
| Cdc42 Pull-down Activation Assay Biochem Kit (bead pull-down format) |
CYT | BK034-S | 1 KIT [20 assays] |
¥132,000 |
| RhoA G-LISA(R) Activation Assay (absorbance) |
CYT | BK124 | 96 ASSAY |
CYT社 BK124L 1 を参照 |
| Rac1 G-LISA(R) Activation Assay Kit |
CYT | BK128 | 1 KIT [96 assays] |
CYT社 BK128L 1 を参照 |
| Cdc42 G-LISA(R) Activation Assay Kit |
CYT | BK127 | 96 ASSAY |
CYT社 BK127L 1 を参照 |
■ CYTOSKELETON NEWS バックナンバー
- 2020年10月号 紡錘体 - 可視化に向けた新規ツール
- 2020年8月号 細胞膜染色用蛍光プローブ
- 2020年4月号 生細胞におけるF-アクチンプローブ
- 2020年3月号 コロナウイルスと細胞骨格
- 2020年2月号 タウ(Tau)の将来性をMapping
- 2020年1月号 Rho GTPaseによる細胞遊走制御
- 2019年12月号 表現型プロファイリング:アクチンに焦点を当てたがん治療
- 2019年11月号 チューブリンの過剰グルタミル化、ミトコンドリア、神経変性
- 2019年9月号 細胞運動性を制御するために相互作用する膜張力とアクチン細胞骨格
- 2019年8月号 Rac1B、がん、およびRac1
- 2019年7月号 Rhoファミリー GTPases、神経可塑性、およびうつ状態
- 2019年6月号 アクチンメチオニン酸化: 動的制御の次の段階
- 2019年5月号 ミクログリアと神経変性疾患
- 2019年2月号 生細胞画像化に対するCNS疾患や障害
2018年
- 2018年12月号 アクチン細胞骨格とメカノトランスダクション(機械的シグナル伝達)
- 2018年11月号 軸索再生と細胞骨格
- 2018年10月号 ニューロンにおける微小管と極性
- 2018年8月号 Rab GTPase と 神経変性
- 2018年7月号 SUMO レスリング: バランスが全て
- 2018年6月号 なぜ K-Ras は発がん特異性を示すのか?
- 2018年5月号 治療標的としてのユビキチンプロテアソームシステム:チューブリンは関与するか?
- 2018年4月号 RhoファミリーGEFと樹状突起スパインの構造的可塑性
- 2018年3月号 βカテニンとTFC/LEF-1の翻訳後修飾による標準的なWntシグナル制御
- 2018年2月号 がん抑制遺伝子p53の翻訳後修飾による機能の調整
- 2018年1月号 自閉スペクトラム症におけるGEF Trioの役割
2017年
- 2017年12月号 プロフィリン: アクチン結合タンパク質の多機能な役割
- 2017年11月号 ミトコンドリアにおけるアセチル化:新たな考え方と治療への応用の可能性
- 2017年9月号 翻訳後修飾のアセチル化による微小管の安定化
- 2017年8月号 神経軸索におけるアクチンリングを基盤とした周期的膜骨格(PMS)
- 2017年7月号 E3ユビキチンリガーゼMdm2によるがん抑制遺伝子p53の翻訳後制御
- 2017年6月号 多能性幹細胞(PSC)での転写因子による翻訳後制御
- 2017年5月号 Arf6 GEF と癌細胞の浸潤・転移
- 2017年4月号 PTEN(Phosphatase and Tensin Homolog)による翻訳後制御
- 2017年3月号 Tau の翻訳後修飾: アルツハイマー病の治療標的
- 2017年2月号 樹状細胞の移動におけるアクチン結合タンパク質とF-アクチン
2016年
- 2016年11月/12月号 GEF を介した GTPase シグナル伝達の低分子阻害剤
- 2016年9月号 FtsZ タンパク質: 抗菌薬の新規ターゲット
- 2016年7月号 翻訳後修飾(PTM)は心臓病において細胞骨格タンパク質を調節する
- 2016年6月号 モータータンパク質キネシンと神経変性
- 2016年5月号 チロシンリン酸化は Rhoファミリー GTPase 活性を調節する
- 2016年4月号 Rac1と糖尿病: ポジティブな役割とネガティブな役割
- 2016年3月号 SUMO化: 細胞骨格タンパク質の機能を調節するレギュレーター
- 2016年1月/2月号 ビメンチン中間径フィラメント: リン酸化による調節
2015年
- 2015年8月号 タンパク質調節に不可欠な翻訳後修飾
- 2015年7月号 アクチン細胞骨格のライブセルイメージング
- 2015年6月号 有糸分裂に関わるタンパク質のSUMO化: 局在と機能
- 2015年5月号 Ras 癌の治療: 5つの有望なターゲット
- 2015年4月号 Ras 依存性の癌で注目される YAP1
- 2015年3月号 増刊号 統合失調症において遺伝子変異により誘導されるアクチン依存のシナプスの変化
- 2015年3月号 Ral GTPase を調節する翻訳後修飾
- 2015年1月/2月号 RhoA のリン酸化はシグナル伝達を調節する
- 2015年1月号 増刊号 微小管を不安定化する suprafenacine: 新規抗癌剤のリード化合物としての可能性
2014年
- 2014年12月号 増刊号 RhoA は心筋細胞におけるアクチン細胞骨格の再構成とグルコース取り込みを仲介する
- 2014年11月号 増刊号 樹状突起の形態形成: ドーパミンD1受容体 および Rho ファミリー GTPase による制御
- 2014年11月/12月号 GTPase 活性化アッセイ: アイソフォームの検出
- 2014年10月号 アルギニンの正電荷を消失させるシトルリン化
- 2014年9月号 キネシンサブドメインの探索
- 2014年9月号 増刊号 アクチン結合タンパク質コフィリンの S-ニトロシル化: 細胞移動に対する影響
- 2014年8月号 増刊号 原発性硬化性胆管炎における N-Ras 発現および活性
- 2014年8月号 SUMO化: 細胞骨格タンパク質を標的とした翻訳後修飾
- 2014年7月号 Sos/K-Ras 結合を介して Ras シグナル伝達を制御する新しい低分子阻害剤
- 2014年6月号 増刊号 頭頸部扁平上皮癌における microRNA-138 による RhoC のダウンレギュレーション
- 2014年6月号 Rho GTPase と活性酸素種: クロストークとフィードバック
- 2014年5月号 ミオシンのアセチル化はサルコメアの構造と機能を調節する
- 2014年4月号 リジンのアセチル化 - 多様な細胞プロセスの制御因子
- 2014年3月号 インテグリンを介したβ-アクチンの酸化還元制御: PDIの出現
- 2014年1/2月号 ダイニン: 一つのモーターが関わる複数の神経変性疾患
2013年
- 2013年11/12月号 ダイニン:チームとして強力に作用するモータータンパク質
- 2013年10月号 神経変性:Rhes、SUMO化、ハンチントン病
- 2013年9月号 モノユビキチン化:タンパク質調節のダイナミックなタグ
- 2013年8月号 Ras及びRhoのプレニル化による翻訳後修飾:癌創薬における役割
- 2013年7月号 アクチンが引き起こす膜突起による浸潤:コルタクチン
- 2013年6月号 アクチン修飾と細胞骨格
- 2013年5月号 微小管内部の実体
- 2013年4月号 神経変性におけるTauの多面性
- 2013年3月号 蛍光フィブロネクチンタンパク質を用いた特発性肺線維症の創薬
- 2013年1/2月号 樹状突起棘:発生におけるArf6の役割
2012年
- 2012年11/12月号 ミオシンの小分子モジュレーター
- 2012年10月号 Rhoファミリーパスウェイのユビキチン化と制御
- 2012年9月号 神経変性におけるRac1 GTPaseの機能
- 2012年8月号 上皮間葉転換(EMT)とRhoファミリー低分子量G-タンパク質の関与
- 2012年7月号 チューブリンの多重修飾:グルタミル化とグリシル化
- 2012年6月号 細胞接着のフィブロネクチン制御と原線維形成
- 2012年5月号 アクチン酸化サイクルの機能
- 2012年4月号 トラフィッキング:ArfとCdc42/Racの結合
- 2012年3月号 G-LISAを用いた心臓研究: 糖尿病性心筋症におけるRho経路に関する研究
- 2012年1月/2月号 FtsZ: 新たな抗生剤の標的となるチュ−ブリンホモログ
商品は「研究用試薬」です。人や動物の医療用・臨床診断用・食品用としては使用しないように、十分ご注意ください。
※ 表示価格について














