






















 このページを印刷する
このページを印刷する
記事ID : 12555
インテグリンを介したβ-アクチンの酸化還元制御: PDIの出現 CYTOSKELETON NEWS 2014年3月号
インテグリンを介したβ-アクチンの酸化還元制御: PDIの出現
細胞運動は、血管新生、創傷治癒、免疫監視機構などの生物学的プロセスにとって不可欠です1。インテグリンは、αβヘテロ二量体膜貫通型受容体で、細胞と細胞外マトリックス(ECM)との細胞基質接着および細胞-細胞の接着に関連しています(図1)。インテグリンが結合するECMタンパク質には、フィブロネクチン、コラーゲン、ラミニンなどがあり、各ECMタンパク質に対して親和性が異なる複数のインテグリンが存在します(例: 9種類のインテグリンアイソフォームがフィブロネクチンに結合できます)2。従って、細胞接着のダイナミクスと運動性は、細胞に発現するインテグリンのレパートリーに影響を受けており2、様々ながんにおけるインテグリンの発現プロファイルは、浸潤性や転移の増大に関連しています3,4。
アクチン細胞骨格へのインテグリン受容体シグナルの伝達は、複雑で多面的なプロセスです(図1)。インテグリン受容体は、ECMに結合してクラスターを形成し、アダプタータンパク質(例: タリン、ビンキュリン、α-アクチニン)、Arp2/3 複合体のリクルート、キナーゼの活性化(例: FAK、ILK など)、低分子量Gタンパク質(例: Rho GTPase)など、複数の方法を介してアクチンにシグナルを伝えます1。また、インテグリンの結合により、さまざまな方法(例: NADPHオキシダーゼ、5-lipoxygenase、ミトコンドリア5)で活性酸素種(ROS)が産生され、アクチンの酸化が起こります。β-アクチンには保存されたシステイン残基が6つありますが、そのうちのCys374 だけが、酸化還元感受性があり、球状(単量体)とフィラメント状(重合体)アクチン(それぞれG-アクチンとF-アクチン)両方で利用できます6-8。β-アクチンの酸化は、インテグリンとフィブロネクチンの結合に応答した細胞の伸展に必要であり、S-グルタチオン化として知られる可逆的なプロセスにおいて、Cys374 とグルタチオン間のジスルフィド結合の形成を介して起こることが示されています9。実際には、この修飾によってアクチンを不可逆的な酸化から保護し、下流で起こるイベントのシグナルとなる、二重の役割があります。β-アクチンのS-グルタチオン化によって起こる重要な結果の一つは、細胞接着や移動に必要なステップであるアクトミオシン複合体(すなわち、F-アクチンとミオシン)の分解です9(図1)。
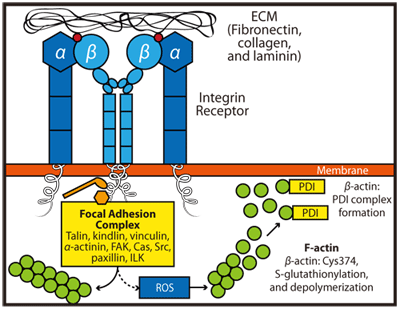
図1 インテグリン受容体とECMの相互作用がアクチンダイナミクスに及ぼす影響。インテグリン受容体とECMタンパク質(フィブロネクチン、コラーゲン、ラミニンなど)の結合により、受容体のクラスター化、接着班複合体の形成、局所的なアクチン重合刺激が起こります。これとは対照的に、下流のシグナル伝達により、β-アクチン Cys374 の酸化を促進する ROS の産生が起こります。その結果、この残基はS-グルタチオン化され、β-アクチンがPDIと会合します。このステップは細胞運動に不可欠です。
S-グルタチオン化からの回復には、酵素に依存する方法と、依存しない方法の両方があります7,10。このプロセスに関与する主要な酵素は、グルタレドキシンだと考えられていますが、チオレドキシン、スルフィレドキシン、タンパク質ジスルフィドイソメラーゼ(PDI)にも、この機能があります7,11。 PDI の標準的な役割は、小胞体(ER)内で新生タンパク質の正確なジスルフィド結合を形成することです12。しかし、PDIは、血小板凝集の際には細胞外でも機能し13,14、シグナル依存的な方法でERから放出されることが示されています15。近年の研究から、PDIは、αIIbβ3インテグリンとフィブロネクチンの結合に応答して起こるβ-アクチンの酸化の調節に、主要な役割を果たしていることが示唆されています16。 Sobierajska ら16 による研究では、PDIは、インテグリンが活性化された状態でβ-アクチン Cys374 と混合ジスルフィド結合を形成することが示されました(図1)。PDIがERの外側で機能することと一致するように、PDIとβ-アクチンが細胞の伸展に関連する膜突起(葉状仮足)に共局在することが、免疫蛍光実験によって示されています。また、β-アクチンとPDI複合体の生理的な関連性については、PDIのアンチセンスRNAを使用してPDI発現を低下させると細胞接着が阻害され、フィブロネクチン上に分布することからも証明されています。
インテグリンシグナル伝達に応答して起こるβ-アクチンの酸化の調節において、PDIが重要な役割を果たしているという発見から、β-アクチンがS-グルタチオン化される他のシグナル伝達経路においても、PDIが重要であることが示唆されます。また、この分野のさらなる研究が、PDIの他のS-グルタチオン化されたタンパク質における幅広い役割を明らかにし、PDIが結合したタンパク質が疾患のバイオマーカーとなることが期待されます。
Cytoskeleton社では、β-アクチンの酸化還元制御研究にご使用いただける、様々なアクチンベースのツールを提供しています。精製β-アクチン(非筋: 非標識およびローダミン標識)、アクチン重合アッセイキット、G-/F-アクチン in vivo アッセイキット、各種アクチン結合タンパク質(例: α-アクチニン、Arp2/3複合体)、抗体などの商品をご用意しています。
参考文献
1. Srichai M.B. and Zent R. 2010. Integrin structure and function. In: Cell-Extracellular Matrix Interactions in Cancer, R. Zent and A. Pozzi, Eds., pp. 19–41, Springer Science+Business Media, LLC., New York, NY.
2. Huttenlocher A. and Horwitz A.R. 2011. Integrins in Cell Migration. Cold Spring Harb. Perspect. Biol. 3, a005074.
3. Seftor R.E.B. et al. 1992. Role of avb3 integrin in human melanoma cell invasion. Proc. Natl. Acad. Sci. U.S.A. 89, 1557-1561.
4. Chan B.M.C. et al. 1990. In vitro and in vivo consequences of VLA-1 expression on rhabdomyosarcoma cells. Science. 251, 1600-1602.
5. Zeller K.S. et al. 2013. The role of mechanical force and ROS in integrin-dependent signals. PLoS ONE. 8, e64897.
6. Stournaras C. 1990. Exposure of thiol groups and bound nucleotide in G-actin: thiols as an indicator for the native state of actin. Anti-Cancer Res. 10, 1651-1659.
7. Dalle-Donne I. et al. 2003. Reversible S-glutathionylation of C374 regulates actin filament formation by inducing structural changes in the actin molecule. Free Radic. Biol. Med. 34, 23-32.
8. Orlova A. et al. 2001. Probing the structure of F-actin: cross-links constrain atomic models and modify actin dynamics. J. Mol. Biol. 312, 95-106.
9. Fiaschi T. et al. 2006. Redox regulation of beta-actin during integrin-mediated cell adhesion. J. Biol. Chem. 281, 22983–22991.
10. Grek C.L. et al. 2013. Causes and consequences of cysteine S-glutathionylation. J. Biol. Chem. 288, 26497-26504.
11. Reinhardt C. et al. 2008. Protein disulfide isomerase acts as an injury response signal that enhances fibrin generation via tissue factor activation. J. Clin. Invest. 118, 1110-1122.
12. Noiva R. 1999. Protein disulfide isomerase: The multifunctional redox chaperone of the endoplasmic reticulum. Semin. Cell Dev. Biol. 10, 481-493.
13. Essex D.W. and Li M. 2003. Redox control of platelet aggregation. Biochemistry. 42, 129-136.
14. Jasuja R. et al. 2010. Endothelium-derived but not platelet-derived protein disulfide isomerase is required for thrombus formation in vivo. Blood. 116, 4665-4674.
15. Safran M. et al. 1992. Thyroid hormone-dependent redistribution of the 55-kilodalton monomer of protein disulfide isomerase in cultured glial cells. Endocrinology. 131, 2413-2418.
16. Sobierajska K. et al. 2014. PDI directly interacts with b-actin Cys374 and regulates cytoskeleton reorganization. J. Biol. Chem. doi: 10.1074/jbc.M113.479477.
アクチン結合タンパク質
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
Acti-stain 488 phalloidin, Plant |
CYT | PHDG1-A | 1*500 UL [300 slides] |
¥79,000 |
| Acti-stain 555 phalloidin, Mushroom |
CYT | PHDH1-A | 1*500 UL [300 slides] |
¥79,000 |
| Acti-stain 670 phalloidin, Plant |
CYT | PHDN1-A | 1*500 UL [300 slides] |
¥79,000 |
| Alpha-actinin, Rabbit, Unlabeled |
CYT | AT01-A | 2*50 UG |
¥110,000 |
| Alpha-actinin, Rabbit, Unlabeled |
CYT | AT01-C | 10*50 UG |
¥368,000 |
| Arp2/3 protein complex, Porcine |
CYT | RP01P-A | 2*50 UG |
¥147,000 |
| Arp2/3 protein complex, Porcine |
CYT | RP01P-B | 6*50 UG |
¥380,000 |
| Cofilin 1, Human, Unlabeled |
CYT | CF01-A | 1*100 UG |
¥91,000 |
| Cofilin 1, Human, Unlabeled |
CYT | CF01-C | 4*100 UG |
¥288,000 |
| Gelsolin, Human |
CYT | HPG6-A | 4*20 UG |
¥123,000 |
| Gelsolin, Human |
CYT | HPG6-B | 20*20 UG |
¥399,000 |
| Heavy meromyosin, Rabbit, Unlabeled |
CYT | MH01-A | 4*50 UG |
¥102,000 |
| Myosin II, Rabbit, Unlabeled |
CYT | MY02-B | 20*1 MG |
¥335,000 |
| Myosin II, Rabbit, Unlabeled |
CYT | MY02-A | 5*1 MG |
¥116,000 |
| Myosin, Bovine |
CYT | MY03-A | 5*1 MG |
¥116,000 |
| Myosin, Bovine |
CYT | MY03-B | 20*1 MG |
¥335,000 |
| Profilin 1, Human, Unlabeled |
CYT | PR01-A | 1*50 UG |
販売終了 |
| Rhodamine Phalloidin, Rhodamine Isothiocyanate |
CYT | PHDR1 | 1*500 UL [300 slides] |
¥79,000 |
| WASP, Human |
CYT | VCG03-A | 1*500 UG |
¥177,000 |
アクチンタンパク質
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
| Actin, Human, Unlabeled |
CYT | APHL99-A | 2*250 UG |
¥79,000 |
| Actin, Human, Unlabeled |
CYT | APHL99-C | 1*1 MG |
¥133,000 |
| Actin, Rabbit, Unlabeled |
CYT | AKL95-B | 1*1 MG |
¥35,000 |
| Actin, Rabbit, Unlabeled |
CYT | AKL95-C | 5*1 MG |
¥137,000 |
| Actin, Bovine, Unlabeled |
CYT | AD99-A | 1*1 MG |
¥44,000 |
| Actin, Bovine, Unlabeled |
CYT | AD99-B | 5*1 MG |
¥181,000 |
| Actin, Chicken |
CYT | AS99-A | 1*1 MG |
¥44,000 |
| Actin, Chicken |
CYT | AS99-B | 5*1 MG |
¥181,000 |
| Actin, Human, Unlabeled |
CYT | APHL99-E | 5*1 MG |
お問い合わせ |
| Actin, Rabbit, Unlabeled |
CYT | AKL99-A | 4*250 UG |
¥73,000 |
| Actin, Rabbit, Unlabeled |
CYT | AKL99-B | 2*1 MG |
¥87,000 |
| Actin, Rabbit, Unlabeled |
CYT | AKL99-C | 5*1 MG |
¥177,000 |
| Actin, Rabbit, Unlabeled |
CYT | AKL99-D | 10*1 MG |
¥335,000 |
| Actin, Rabbit, Unlabeled |
CYT | AKL99-E | 20*1 MG |
お問い合わせ |
| Actin, Rabbit, Biotin |
CYT | AB07-A | 5*20 UG |
¥120,000 |
| Actin, Rabbit, Biotin |
CYT | AB07-C | 20*20 UG |
¥344,000 |
| Actin protein (pre-formed filaments), Rabbit |
CYT | AKF99-A | 1*1 MG |
¥69,000 |
| Actin protein (pre-formed filaments), Rabbit |
CYT | AKF99-B | 5*1 MG |
¥240,000 |
| Actin, Rabbit, Pyrene |
CYT | AP05-A | 1*1 MG |
¥75,000 |
| Actin, Rabbit, Pyrene |
CYT | AP05-B | 5*1 MG |
¥342,000 |
| Actin, Human, Rhodamine Isothiocyanate |
CYT | APHR-A | 4*10 UG |
¥108,000 |
| Actin, Human, Rhodamine Isothiocyanate |
CYT | APHR-C | 20*10 UG |
¥339,000 |
| Actin, Rabbit, Rhodamine Isothiocyanate |
CYT | AR05-B | 10*20 UG |
¥104,000 |
| Actin, Rabbit, Rhodamine Isothiocyanate |
CYT | AR05-C | 20*20 UG |
¥200,000 |
アクチン関連タンパク質
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
| Anti Cofilin w/positive control, Human (Rabbit) Unlabeled |
CYT | ACFL02-A | 1*50 UG |
販売終了 |
| Anti Cofilin w/positive control, Human (Rabbit) Unlabeled |
CYT | ACFL02-B | 3*50 UG |
販売終了 |
| Anti Actin w/positive control, Human (Rabbit) Unlabeled |
CYT | AAN01-A | 1*100 UG |
販売終了 |
| Anti Actin w/positive control, Human (Rabbit) Unlabeled |
CYT | AAN01-B | 3*100 UG |
販売終了 |
| Anti Profilin w/positive control, Human (Rabbit) Unlabeled |
CYT | APUF01-A | 1*50 UG |
販売終了 |
アクチンタンパク質キット
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
| Actin Binding Protein Spin-Down Assay Biochem Kit, Human |
CYT | BK013 | 1 KIT [30-100 assays] |
¥272,000 |
| Actin Binding Protein Spin-Down Assay Biochem Kit, Rabbit |
CYT | BK001 | 1 KIT [30-100 assays] |
¥268,000 |
| Actin Polymerization Biochem Kit (fluorescence format), Rabbit |
CYT | BK003 | 1 KIT [30-100 assays] |
¥304,000 |
| F-actin Visualization Biochem Kit (fluorescence format) |
CYT | BK005 | 1 KIT [300 slides] |
¥141,000 |
| G-Actin/F-actin In Vivo Assay Biochem Kit, Mouse |
CYT | BK037 | 1 KIT [30-100 assays] |
¥264,000 |
■ CYTOSKELETON NEWS バックナンバー
- 2020年10月号 紡錘体 - 可視化に向けた新規ツール
- 2020年8月号 細胞膜染色用蛍光プローブ
- 2020年4月号 生細胞におけるF-アクチンプローブ
- 2020年3月号 コロナウイルスと細胞骨格
- 2020年2月号 タウ(Tau)の将来性をMapping
- 2020年1月号 Rho GTPaseによる細胞遊走制御
- 2019年12月号 表現型プロファイリング:アクチンに焦点を当てたがん治療
- 2019年11月号 チューブリンの過剰グルタミル化、ミトコンドリア、神経変性
- 2019年9月号 細胞運動性を制御するために相互作用する膜張力とアクチン細胞骨格
- 2019年8月号 Rac1B、がん、およびRac1
- 2019年7月号 Rhoファミリー GTPases、神経可塑性、およびうつ状態
- 2019年6月号 アクチンメチオニン酸化: 動的制御の次の段階
- 2019年5月号 ミクログリアと神経変性疾患
- 2019年2月号 生細胞画像化に対するCNS疾患や障害
2018年
- 2018年12月号 アクチン細胞骨格とメカノトランスダクション(機械的シグナル伝達)
- 2018年11月号 軸索再生と細胞骨格
- 2018年10月号 ニューロンにおける微小管と極性
- 2018年8月号 Rab GTPase と 神経変性
- 2018年7月号 SUMO レスリング: バランスが全て
- 2018年6月号 なぜ K-Ras は発がん特異性を示すのか?
- 2018年5月号 治療標的としてのユビキチンプロテアソームシステム:チューブリンは関与するか?
- 2018年4月号 RhoファミリーGEFと樹状突起スパインの構造的可塑性
- 2018年3月号 βカテニンとTFC/LEF-1の翻訳後修飾による標準的なWntシグナル制御
- 2018年2月号 がん抑制遺伝子p53の翻訳後修飾による機能の調整
- 2018年1月号 自閉スペクトラム症におけるGEF Trioの役割
2017年
- 2017年12月号 プロフィリン: アクチン結合タンパク質の多機能な役割
- 2017年11月号 ミトコンドリアにおけるアセチル化:新たな考え方と治療への応用の可能性
- 2017年9月号 翻訳後修飾のアセチル化による微小管の安定化
- 2017年8月号 神経軸索におけるアクチンリングを基盤とした周期的膜骨格(PMS)
- 2017年7月号 E3ユビキチンリガーゼMdm2によるがん抑制遺伝子p53の翻訳後制御
- 2017年6月号 多能性幹細胞(PSC)での転写因子による翻訳後制御
- 2017年5月号 Arf6 GEF と癌細胞の浸潤・転移
- 2017年4月号 PTEN(Phosphatase and Tensin Homolog)による翻訳後制御
- 2017年3月号 Tau の翻訳後修飾: アルツハイマー病の治療標的
- 2017年2月号 樹状細胞の移動におけるアクチン結合タンパク質とF-アクチン
2016年
- 2016年11月/12月号 GEF を介した GTPase シグナル伝達の低分子阻害剤
- 2016年9月号 FtsZ タンパク質: 抗菌薬の新規ターゲット
- 2016年7月号 翻訳後修飾(PTM)は心臓病において細胞骨格タンパク質を調節する
- 2016年6月号 モータータンパク質キネシンと神経変性
- 2016年5月号 チロシンリン酸化は Rhoファミリー GTPase 活性を調節する
- 2016年4月号 Rac1と糖尿病: ポジティブな役割とネガティブな役割
- 2016年3月号 SUMO化: 細胞骨格タンパク質の機能を調節するレギュレーター
- 2016年1月/2月号 ビメンチン中間径フィラメント: リン酸化による調節
2015年
- 2015年8月号 タンパク質調節に不可欠な翻訳後修飾
- 2015年7月号 アクチン細胞骨格のライブセルイメージング
- 2015年6月号 有糸分裂に関わるタンパク質のSUMO化: 局在と機能
- 2015年5月号 Ras 癌の治療: 5つの有望なターゲット
- 2015年4月号 Ras 依存性の癌で注目される YAP1
- 2015年3月号 増刊号 統合失調症において遺伝子変異により誘導されるアクチン依存のシナプスの変化
- 2015年3月号 Ral GTPase を調節する翻訳後修飾
- 2015年1月/2月号 RhoA のリン酸化はシグナル伝達を調節する
- 2015年1月号 増刊号 微小管を不安定化する suprafenacine: 新規抗癌剤のリード化合物としての可能性
2014年
- 2014年12月号 増刊号 RhoA は心筋細胞におけるアクチン細胞骨格の再構成とグルコース取り込みを仲介する
- 2014年11月号 増刊号 樹状突起の形態形成: ドーパミンD1受容体 および Rho ファミリー GTPase による制御
- 2014年11月/12月号 GTPase 活性化アッセイ: アイソフォームの検出
- 2014年10月号 アルギニンの正電荷を消失させるシトルリン化
- 2014年9月号 キネシンサブドメインの探索
- 2014年9月号 増刊号 アクチン結合タンパク質コフィリンの S-ニトロシル化: 細胞移動に対する影響
- 2014年8月号 増刊号 原発性硬化性胆管炎における N-Ras 発現および活性
- 2014年8月号 SUMO化: 細胞骨格タンパク質を標的とした翻訳後修飾
- 2014年7月号 Sos/K-Ras 結合を介して Ras シグナル伝達を制御する新しい低分子阻害剤
- 2014年6月号 増刊号 頭頸部扁平上皮癌における microRNA-138 による RhoC のダウンレギュレーション
- 2014年6月号 Rho GTPase と活性酸素種: クロストークとフィードバック
- 2014年5月号 ミオシンのアセチル化はサルコメアの構造と機能を調節する
- 2014年4月号 リジンのアセチル化 - 多様な細胞プロセスの制御因子
- 2014年3月号 インテグリンを介したβ-アクチンの酸化還元制御: PDIの出現
- 2014年1/2月号 ダイニン: 一つのモーターが関わる複数の神経変性疾患
2013年
- 2013年11/12月号 ダイニン:チームとして強力に作用するモータータンパク質
- 2013年10月号 神経変性:Rhes、SUMO化、ハンチントン病
- 2013年9月号 モノユビキチン化:タンパク質調節のダイナミックなタグ
- 2013年8月号 Ras及びRhoのプレニル化による翻訳後修飾:癌創薬における役割
- 2013年7月号 アクチンが引き起こす膜突起による浸潤:コルタクチン
- 2013年6月号 アクチン修飾と細胞骨格
- 2013年5月号 微小管内部の実体
- 2013年4月号 神経変性におけるTauの多面性
- 2013年3月号 蛍光フィブロネクチンタンパク質を用いた特発性肺線維症の創薬
- 2013年1/2月号 樹状突起棘:発生におけるArf6の役割
2012年
- 2012年11/12月号 ミオシンの小分子モジュレーター
- 2012年10月号 Rhoファミリーパスウェイのユビキチン化と制御
- 2012年9月号 神経変性におけるRac1 GTPaseの機能
- 2012年8月号 上皮間葉転換(EMT)とRhoファミリー低分子量G-タンパク質の関与
- 2012年7月号 チューブリンの多重修飾:グルタミル化とグリシル化
- 2012年6月号 細胞接着のフィブロネクチン制御と原線維形成
- 2012年5月号 アクチン酸化サイクルの機能
- 2012年4月号 トラフィッキング:ArfとCdc42/Racの結合
- 2012年3月号 G-LISAを用いた心臓研究: 糖尿病性心筋症におけるRho経路に関する研究
- 2012年1月/2月号 FtsZ: 新たな抗生剤の標的となるチュ−ブリンホモログ
商品は「研究用試薬」です。人や動物の医療用・臨床診断用・食品用としては使用しないように、十分ご注意ください。
※ 表示価格について














