






















 このページを印刷する
このページを印刷する
記事ID : 33336
神経軸索におけるアクチンリングを基盤とした周期的膜骨格(PMS) CYTOSKELETON NEWS 2017年8月号
神経軸索におけるアクチンリングを基盤とした周期的膜骨格 (PMS)
アクチンは、神経細胞極性、細胞形態、神経突起の発生(例えば、葉状仮足および糸状仮足の伸長、ならびに樹状突起スパインをもつ成長円錐)、細胞内輸送、およびシナプス可塑性(樹状突起棘数や形態の動的変化 )の制御に関与することから、神経細胞骨格の不可欠な部分です1-3。アクチンが成長円錐や樹状突起棘に存在することが何十年にもわたり研究者に注目されてきた一方で、神経軸索にも存在することで "神経アクチンファミリーの異端児"として取り扱われてきました4。なぜなら、軸索におけるアクチンの構造および役割の正確な詳細がよく分かっていないからです。近年、ナノスケール顕微鏡による周期的膜骨格(PMS)の発見によって、軸索アクチンの構造が解き明かされるという著しい進展がありました(図1)5。本稿では、軸索におけるPMSの発見、構造、および機能に関して論じていきます。
PMSの発見と構造
2013年に発見されたPMSは、皮質アクチンの一種で、アクチン皮質の主要な構成成分です。PMSは、F-アクチンとアクチン結合タンパク質の混合物で、エンドサイトーシス/エキソサイトーシスおよび細胞運動性といった真核生物細胞の細胞膜とそれに関するプロセスを担っています4, 5。初節を含めた神経軸索では6、短鎖アクチンフィラメントによって構成されるPMSが等間隔の輪に束ねられ、180〜190 nm の周期性で軸索の円周に巻かれて構成されています5-9(図1)。短鎖フィラメントはアデューシンキャップにより安定化されており、アクチンリングと軸索の直径を調節するとともに、リング内のアクチンフィラメント成長を調節しています6, 10。アクチンリングの近隣はスペクトリン四量体(軸索本体ではβII、軸索初節ではβIV)による架橋反応で守られています6,8,11。
固定化された哺乳動物神経と脳組織切片に対して、STORM顕微鏡法(stochastic optical reconstruction microscopy)によって、PMSが初めて報告されました5。これらの発見はそのすぐ後、他の研究者により、哺乳動物/非哺乳動物の固定化された神経について、STORM顕微鏡、および SIM顕微鏡法(structured illumination microscopy)を用いて確認されました5,6,8,11,12。最も重要なことに、F-アクチン生細胞イメージングプローブ (SiR-Actin)7,10,13(図1: シリコンローダミン アクチン)を用いて、または蛍光標識済みβIIスペクトリンの外因性発現を行って、生きた哺乳動物の神経を確認したことにより、確証が得られました8。
哺乳動物の培養神経では、発生から最初の数日間で、PMSは神経発達に伴い軸索にそって遠位に移動する前に、細胞体に隣接している軸索内に構築されます5, 7, 8, 11。ショウジョウバエでの一次ニューロンでは、播種から数時間以内でPMS発生が始まります11。特に、ショウジョウバエ神経のPMSについて、非常に幼いもの(数時間から2日齢)と成熟したもの(3日齢以上)との違いが報告されています。例えば、幼い時期のPMSでは軸索成長はアクチン重合(重合核形成因子など)に依存しますが、成熟したものはこの限りではありません11 。重合核形成、集合、およびアクチンリングの維持に関与するアクチン結合タンパク質はあまりよく知られていません。ショウジョウバエ神経では、2つの核形成因子であるArp2/3とホルミンDAAM (disheveled-associated activator of morphogenesis)がPMS形成をし、おそらくPMSを構成する複数の短鎖フィラメントが重合核となっていると思われます11。
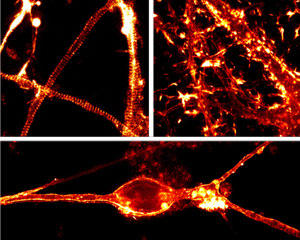
図1. STED顕微鏡と100 nM SiR-Actinを用いた神経における生細胞画像
上図左:ラットの海馬神経細胞、8 DIV
上図右:ラットの海馬神経細胞、17 DIV
下図:ラットの小脳顆粒細胞、21 DIV
(マックス・プランク生物物理化学研究所(ゲッチンゲン), Elisa D'Este氏提供)
PMSの機能
神経軸索は、神経が細胞体と軸索終末間を双方向に積荷を伝達し移送する手段です。軸索欠損は永久的なもので、通常の老化・様々な神経変性疾患や病気によって軸索が損なわれることから、正常な脳機能には健康な軸索を維持することが必要です14。PMSは軸索の細胞膜の骨組みとして機能するとの仮説が立てられています。PMSは、アンキリンやナトリウムチャネルのような、それぞれ軸索構造や活動電位発生に重要な分子を、軸索において周期性分布をもつように配置したり5,8、特に軸索末梢部分に沿ってモーター結合積荷小胞のドッキングを補助したり4,9,15、軸索に沿って膜貫通型タンパク質を形成したりしています11, 15-18 。
実際の機能データはわずかですが、固定化し、SiR-Actinで染色したショウジョウバエ一次ニューロンに関する最近の報告では、軸索の完全性を維持する上で、PMSは軸索微小管(MT)の重合を促進することから重要であると結論づけられています11。軸索微小管は、軸索における主要な細胞骨格成分です。サイトカラシンDに誘導させてPMSを分解すると、微小管束の切断、微小管重合の低減、軸索数の減少が見られます11。これ以外の微小管に関連するPMSの機能的役割としては、微小管を束ねることが挙げられます19-21。PMSは、微小管のマイナス末端へのアンカーとして機能し22, 23 、ダイニンが微小管上をすべるのを助けたり、移送を補助したりしています24。
まとめ
近年、神経軸索の膜下格子を構成するアクチンリングが発見され、軸索アクチンの役割をより深く理解できるすばらしい機会がありました。さらに、末梢神経系のランビエ絞輪や、少なくともいくつかの樹状突起や樹状突起スパインにおいてもアクチンリングの記述がなされています7, 25。アクチンリングと似ているものに、「アクチンの波」と「アクチンの通り道」というアクチン構造がありますが、あまり注目されてきませんでした4。超解像顕微鏡(STORM, STED, SIMなど)やSiR-Actinといった生細胞画像プローブなしでは、PMSの存在や組成は未知のままだったでしょう。アクチンや他の細胞骨格構造の組成、集合、維持、および機能をさらに明らかにできるよう、Cytoskeleton社では、F-アクチン、微小管、DNA、リソソーム用の生細胞イメージングプローブ (SiRシリーズ、SiR700シリーズ)や、固定化細胞用Acti-stainファロイジンをご提供しています。
参考文献
- Luo L. 2002. Actin cytoskeleton regulation in neuronal morphogenesis and structural plasticity. Annu. Rev. Cell Dev. Biol. 18, 601-635.
- Cingolani L.A. and Goda Y. 2008. Actin in action: The interplay between the actin cytoskeleton and synaptic efficacy. Nat. Rev. Neurosci. 9, 344-356.
- Stiess M. and Bradke F. 2011. Neuronal polarization: The cytoskeleton leads the way. Dev. Neurobiol. 71, 430-444.
- Roy S. 2016. Waves, rings, and trails: The scenic landscape of axonal actin. J. Cell Biol. 212, 131-134.
- Xu K. et al. 2013. Actin, spectrin, and associated proteins form a periodic cytoskeletal structure in axons. Science. 339, 452-456.
- Leterrier C. et al. 2015. Nanoscale architecture of the axon initial segment reveals an organized and robust scaffold. Cell Rep. 13, 2781-2793.
- D’Este E. et al. 2015. STED nanoscopy reveals the ubiquity of subcortical cytoskeleton periodicity in living neurons. Cell Rep. 10, 1246-1251.
- Zhong G. et al. 2014. Developmental mechanism of the periodic membrane skeleton in axons. eLife. 3, e04581.
- Leite S.C. and Sousa M.M. 2016. The neuronal and actin commitment: Why do neurons need rings? Cytoskeleton. 73, 424-434.
- Leite S.C. et al. 2016. The actin-binding protein a-adducin is required for maintaining axon diameter. Cell Rep. 15, 490-498.
- Qu Y. et al. 2017. Periodic actin structures in neuronal axons are required to maintain microtubules. Mol. Biol. Cell. 28, 296-308.
- He J. et al. 2016. Prevalent presence of periodic actin-spectrin-based membrane skeleton in a broad range of neuronal cell types and animal species. Proc. Natl. Acad. Sci. U.S.A. 113, 6029-6034.
- Lukinavicius G. et al. 2014. Fluorogenic probes for live-cell imaging of the cytoskeleton. Nat. Methods. 11, 731-733.
- Adalbert R. and Coleman M.P. 2012. Axon pathology in age-related neurodegenerative disorders. Neuropathol. Appl. Neurobiol. 39, 90-108.
- Gallo G. 2013. More than one ring to bind them all: Recent insights into the structure of the axon. Dev. Neurobiol. 73, 799-805.
- Zhang Y. et al. 2016. Axon membrane skeleton structure is optimized for coordinated sodium propagation. arXiv.1602.06348.
- Albrecht D. et al. 2016. Nanoscopic compartmentalization of membrane protein motion at the axon initial segment. J. Cell Biol. 215, 37-46.
- Machnicka B. et al. 2014. Spectrins: a structural platform for stabilization and activation of membrane channels, receptors and transporters. Biochim. Biophys. Acta. 1838, 620-634.
- Prokop A. et al. 2013. Using fly genetics to dissect the cytoskeletal machinery of neurons during axonal growth and maintenance. J. Cell Sci. 126, 2331-2341.
- Sanchez-Soriano N. et al. 2009. Mouse ACF7 and Drosophila Short stop modulate filopodia formation and microtubule organization during neuronal growth. J. Cell Sci. 122, 2534-2542.
- Alves-Silva J. et al. 2012. Spectraplakins promote microtubule-mediated axonal growth by functioning as structural microtubule-associated proteins and EB1-dependent +TIPs (tip interaction proteins). J. Neurosci. 32, 9143-9158.
- Nashchekin D. et al. 2016. Patronin/Shot cortical foci assemble the noncentrosomal microtubule array that specifies the Drosophila anterior-posterior axis. Dev. Cell. 38, 61-72.
- Ning W. et al. 2016. The CAMSAP3-ACF7 complex couples noncentrosomal microtubules with actin filaments to coordinate their dynamics. Dev. Cell. 39, 61-74.
- Myers K.A. et al. 2006. Microtubule transport in the axon: Re-thinking a potential role for the actin cytoskeleton. Neuroscientist. 12, 107-118.
- Bar J. et al. 2016. Periodic F-actin structures shape the neck of dendritic spines. Sci. Rep. 6, 37136.
Spirochrome™Live Cell Imaging Probes
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
SiR700-Actin Kit |
CYT | CY-SC013 | 1 KIT [35-200 slides] |
¥182,000 |
| SiR-Actin Kit |
CYT | CY-SC001 | 1 KIT [50-300 slides] |
¥182,000 |
| SiR700-Tubulin Kit |
CYT | CY-SC014 | 1 KIT [35-200 slides] |
¥182,000 |
| SiR-Tubulin Kit |
CYT | CY-SC002 | 1 KIT [50-300 slides] |
¥182,000 |
| SiR-Lysosome Kit |
CYT | CY-SC012 | 1 KIT [50-300 slides] |
¥131,000 |
| SiR700-Lysosome Kit |
CYT | CY-SC016 | 1 KIT [35-200 slides] |
¥131,000 |
| SiR-DNA Kit |
CYT | CY-SC007 | 1 KIT [50-300 slides] |
¥120,000 |
| SiR700-DNA Kit |
CYT | CY-SC015 | 1 KIT [35-200 slides] |
¥120,000 |
| Cytoskeleton Kit (SiR-Actin + SiR-Tubulin) |
CYT | CY-SC006 | 1 KIT [50-300 slides] |
販売終了 |
【商品情報】
- Spirochrome プローブ(SiR700-Actin / SiR700-Tubulin / SiR700-DNA)[CYT]
- 細胞骨格(アクチン・チューブリン)やDNAを超解像度で蛍光観察 - Spirochrome プローブキット(SiR-Lysosome / SiR700-Lysosome)[CYT]
- 生細胞中のリソソームを far-red(遠赤色光)でイメージング
Actin Proteins
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
| Actin, Human, Rhodamine Isothiocyanate |
CYT | APHR-A | 4*10 UG |
¥108,000 |
| Actin, Human, Rhodamine Isothiocyanate |
CYT | APHR-C | 20*10 UG |
¥339,000 |
| Actin, Rabbit, Rhodamine Isothiocyanate |
CYT | AR05-B | 10*20 UG |
¥104,000 |
| Actin, Rabbit, Rhodamine Isothiocyanate |
CYT | AR05-C | 20*20 UG |
¥200,000 |
【商品情報】
- アクチンタンパク質・アクチン結合タンパク質[CYT]
- 細胞骨格研究に有用な高品質製品!様々な種類を販売!
Acti-stain Phalloidins
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
| Acti-stain 488 phalloidin, Plant |
CYT | PHDG1-A | 1*500 UL [300 slides] |
¥79,000 |
| Acti-stain 555 phalloidin, Mushroom |
CYT | PHDH1-A | 1*500 UL [300 slides] |
¥79,000 |
| Acti-stain 670 phalloidin, Plant |
CYT | PHDN1-A | 1*500 UL [300 slides] |
¥79,000 |
| Rhodamine Phalloidin, Rhodamine Isothiocyanate |
CYT | PHDR1 | 1*500 UL [300 slides] |
¥79,000 |
【商品情報】
- Acti-Stain™ 蛍光標識ファロイジン[CYT]
- 生細胞/固定細胞の細胞骨格研究に有用
Actin Activation Assay Biochem Kit
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
| G-Actin/F-actin In Vivo Assay Biochem Kit, Mouse |
CYT | BK037 | 1 KIT [30-100 assays] |
¥264,000 |
【商品情報】
- G-アクチン/F-アクチン in vivo Biochem Kit™[CYT]
- G-actin / F-actin ratio を測定するキット
■ CYTOSKELETON NEWS バックナンバー
- 2020年10月号 紡錘体 - 可視化に向けた新規ツール
- 2020年8月号 細胞膜染色用蛍光プローブ
- 2020年4月号 生細胞におけるF-アクチンプローブ
- 2020年3月号 コロナウイルスと細胞骨格
- 2020年2月号 タウ(Tau)の将来性をMapping
- 2020年1月号 Rho GTPaseによる細胞遊走制御
- 2019年12月号 表現型プロファイリング:アクチンに焦点を当てたがん治療
- 2019年11月号 チューブリンの過剰グルタミル化、ミトコンドリア、神経変性
- 2019年9月号 細胞運動性を制御するために相互作用する膜張力とアクチン細胞骨格
- 2019年8月号 Rac1B、がん、およびRac1
- 2019年7月号 Rhoファミリー GTPases、神経可塑性、およびうつ状態
- 2019年6月号 アクチンメチオニン酸化: 動的制御の次の段階
- 2019年5月号 ミクログリアと神経変性疾患
- 2019年2月号 生細胞画像化に対するCNS疾患や障害
2018年
- 2018年12月号 アクチン細胞骨格とメカノトランスダクション(機械的シグナル伝達)
- 2018年11月号 軸索再生と細胞骨格
- 2018年10月号 ニューロンにおける微小管と極性
- 2018年8月号 Rab GTPase と 神経変性
- 2018年7月号 SUMO レスリング: バランスが全て
- 2018年6月号 なぜ K-Ras は発がん特異性を示すのか?
- 2018年5月号 治療標的としてのユビキチンプロテアソームシステム:チューブリンは関与するか?
- 2018年4月号 RhoファミリーGEFと樹状突起スパインの構造的可塑性
- 2018年3月号 βカテニンとTFC/LEF-1の翻訳後修飾による標準的なWntシグナル制御
- 2018年2月号 がん抑制遺伝子p53の翻訳後修飾による機能の調整
- 2018年1月号 自閉スペクトラム症におけるGEF Trioの役割
2017年
- 2017年12月号 プロフィリン: アクチン結合タンパク質の多機能な役割
- 2017年11月号 ミトコンドリアにおけるアセチル化:新たな考え方と治療への応用の可能性
- 2017年9月号 翻訳後修飾のアセチル化による微小管の安定化
- 2017年8月号 神経軸索におけるアクチンリングを基盤とした周期的膜骨格(PMS)
- 2017年7月号 E3ユビキチンリガーゼMdm2によるがん抑制遺伝子p53の翻訳後制御
- 2017年6月号 多能性幹細胞(PSC)での転写因子による翻訳後制御
- 2017年5月号 Arf6 GEF と癌細胞の浸潤・転移
- 2017年4月号 PTEN(Phosphatase and Tensin Homolog)による翻訳後制御
- 2017年3月号 Tau の翻訳後修飾: アルツハイマー病の治療標的
- 2017年2月号 樹状細胞の移動におけるアクチン結合タンパク質とF-アクチン
2016年
- 2016年11月/12月号 GEF を介した GTPase シグナル伝達の低分子阻害剤
- 2016年9月号 FtsZ タンパク質: 抗菌薬の新規ターゲット
- 2016年7月号 翻訳後修飾(PTM)は心臓病において細胞骨格タンパク質を調節する
- 2016年6月号 モータータンパク質キネシンと神経変性
- 2016年5月号 チロシンリン酸化は Rhoファミリー GTPase 活性を調節する
- 2016年4月号 Rac1と糖尿病: ポジティブな役割とネガティブな役割
- 2016年3月号 SUMO化: 細胞骨格タンパク質の機能を調節するレギュレーター
- 2016年1月/2月号 ビメンチン中間径フィラメント: リン酸化による調節
2015年
- 2015年8月号 タンパク質調節に不可欠な翻訳後修飾
- 2015年7月号 アクチン細胞骨格のライブセルイメージング
- 2015年6月号 有糸分裂に関わるタンパク質のSUMO化: 局在と機能
- 2015年5月号 Ras 癌の治療: 5つの有望なターゲット
- 2015年4月号 Ras 依存性の癌で注目される YAP1
- 2015年3月号 増刊号 統合失調症において遺伝子変異により誘導されるアクチン依存のシナプスの変化
- 2015年3月号 Ral GTPase を調節する翻訳後修飾
- 2015年1月/2月号 RhoA のリン酸化はシグナル伝達を調節する
- 2015年1月号 増刊号 微小管を不安定化する suprafenacine: 新規抗癌剤のリード化合物としての可能性
2014年
- 2014年12月号 増刊号 RhoA は心筋細胞におけるアクチン細胞骨格の再構成とグルコース取り込みを仲介する
- 2014年11月号 増刊号 樹状突起の形態形成: ドーパミンD1受容体 および Rho ファミリー GTPase による制御
- 2014年11月/12月号 GTPase 活性化アッセイ: アイソフォームの検出
- 2014年10月号 アルギニンの正電荷を消失させるシトルリン化
- 2014年9月号 キネシンサブドメインの探索
- 2014年9月号 増刊号 アクチン結合タンパク質コフィリンの S-ニトロシル化: 細胞移動に対する影響
- 2014年8月号 増刊号 原発性硬化性胆管炎における N-Ras 発現および活性
- 2014年8月号 SUMO化: 細胞骨格タンパク質を標的とした翻訳後修飾
- 2014年7月号 Sos/K-Ras 結合を介して Ras シグナル伝達を制御する新しい低分子阻害剤
- 2014年6月号 増刊号 頭頸部扁平上皮癌における microRNA-138 による RhoC のダウンレギュレーション
- 2014年6月号 Rho GTPase と活性酸素種: クロストークとフィードバック
- 2014年5月号 ミオシンのアセチル化はサルコメアの構造と機能を調節する
- 2014年4月号 リジンのアセチル化 - 多様な細胞プロセスの制御因子
- 2014年3月号 インテグリンを介したβ-アクチンの酸化還元制御: PDIの出現
- 2014年1/2月号 ダイニン: 一つのモーターが関わる複数の神経変性疾患
2013年
- 2013年11/12月号 ダイニン:チームとして強力に作用するモータータンパク質
- 2013年10月号 神経変性:Rhes、SUMO化、ハンチントン病
- 2013年9月号 モノユビキチン化:タンパク質調節のダイナミックなタグ
- 2013年8月号 Ras及びRhoのプレニル化による翻訳後修飾:癌創薬における役割
- 2013年7月号 アクチンが引き起こす膜突起による浸潤:コルタクチン
- 2013年6月号 アクチン修飾と細胞骨格
- 2013年5月号 微小管内部の実体
- 2013年4月号 神経変性におけるTauの多面性
- 2013年3月号 蛍光フィブロネクチンタンパク質を用いた特発性肺線維症の創薬
- 2013年1/2月号 樹状突起棘:発生におけるArf6の役割
2012年
- 2012年11/12月号 ミオシンの小分子モジュレーター
- 2012年10月号 Rhoファミリーパスウェイのユビキチン化と制御
- 2012年9月号 神経変性におけるRac1 GTPaseの機能
- 2012年8月号 上皮間葉転換(EMT)とRhoファミリー低分子量G-タンパク質の関与
- 2012年7月号 チューブリンの多重修飾:グルタミル化とグリシル化
- 2012年6月号 細胞接着のフィブロネクチン制御と原線維形成
- 2012年5月号 アクチン酸化サイクルの機能
- 2012年4月号 トラフィッキング:ArfとCdc42/Racの結合
- 2012年3月号 G-LISAを用いた心臓研究: 糖尿病性心筋症におけるRho経路に関する研究
- 2012年1月/2月号 FtsZ: 新たな抗生剤の標的となるチュ−ブリンホモログ
商品は「研究用試薬」です。人や動物の医療用・臨床診断用・食品用としては使用しないように、十分ご注意ください。
※ 表示価格について














