






















 このページを印刷する
このページを印刷する
記事ID : 13630
キネシンサブドメインの探索 CYTOSKELETON NEWS 2014年9月号
キネシンサブドメインの探索
近年、分子モーター研究は新しい局面に移りつつあります。巨大分子へのアプローチが中心だった5年以上前とは対照的に1、最近の研究は、キネシンが分子内ダイナミクスによってどのように調節されているか、そのメカニズムの解明にフォーカスされています。主に2つの技術的なアプローチによって、これらの動的なメカニズムの測定が可能になりました。1つは、1分子のキネシンを観察できる全反射照明蛍光(TIRF)顕微鏡で、もう1つは、1つのモーターから別のモーターへのサブドメインスワッピングです。
興味深いことに、キネシンモータータンパク質は、他のキネシンからの小さなアミノ酸ストレッチの置換を、比較的柔軟に受け入れます。キネシンは、通常2つのドメインから構成されています。1つは触媒部位で、もう1つは ネックリンカー(NL)、コイルドコイル(CC)、カーゴ結合部位を含む非触媒性の領域です。NL サブドメインは、長さが 14-22アミノ酸で(図1参照)、Hoeprich ら2 によって、このサブドメインはキネシン1(K-1)からキネシン2(K-2)にスワップが可能で、重要な機能の違いをもスワップして新しい相手先に移行する、と報告されています。Hoeprich ら2 は、1分子のキネシンを追跡することで、MAPs(例: tau)が微小管の表面を覆っている場合、NL サブドメインの長さが、微小管(MT)上を移動する能力を調節していることを示しました、。これらの障害物を横断する際に、K-2 はほとんど失敗しませんが、K-1 はぎこちなく、MAPs で覆われていない MT と比較してよりゆっくりと進みます。モーターが MT から解離する前に移動した距離(processivity または run length)を測定すると、K-2 は、MT が Tau アイソフォーム(3RS または 4RL)で覆われている/いないに関わらず、平均して同じ距離を移動します。一方、K-1 は、Tau で覆われた MT 上をより短い距離しか移動しません。これらのモーター間で NL がスワップされると、特徴的な processivity も一緒にスワップされます。
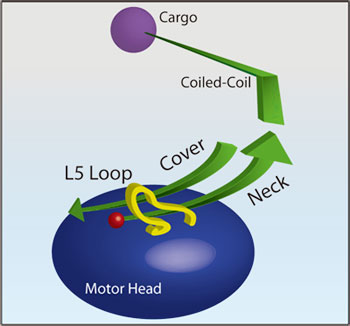
図1 キネシンの模式図
ループL5、ネック、モータードメインに関係するカバーネック、コイルドコイル、カーゴの位置を示す。
互いに近接しており、化学エネルギーを運動エネルギーに変換する際に相互作用することに注目。
Düselder ら3 によって同様の解析が行われ、K-5(Eg5 または KIF11 としても知られている)の processivity の決定に NL の長さが重要であることが報告されました。本研究では、NL が比較的長い(例: 13-22 アミノ酸)場合には、processivity は変わりませんが、ストレッチがより短い(11 アミノ酸)場合には、processivity が低下することが示されています。対照的に、速度および力発生は、NL の長さの変化にそれほど影響されません。しかし、Shastry と Hancock は、processivity に最適なのは 14 アミノ酸である、という異なる結果を報告しています4,5。Shastry と Hancock は、Düselder らと結果が一致しない理由は、使用したバッファーの差異による可能性があり、バッファーは親和性や run length に大きく影響する、と言及しています。興味深いことに、NL 領域の配列と長さはモーターごとに非常に特異的であり、各モーターの機能に最適化されていると考えられます。表1から、NL の長さは 14-22 アミノ酸で、プロリンを含む場合があることがわかります。プロリンは、構造にねじれを形成すると推測され、コイルドコイルをモータードメインの後ろに折り曲げるか、モーターの移動方向を変えることで(例: ホッピングまたは円周方向への移動)、モーター活性の自己阻害を引き起こす可能性があります。
| モーター | キネシンファミリー | NL の前の ID 配列 | NL | NL の後の ID 配列 | NLのアミノ酸長 | processivity (μm) | 参考文献 |
|---|---|---|---|---|---|---|---|
| HsKIF5C (KHC) | K-1 | LDFGRRA | KTVKNVVCVNEELT----- | AEEWKRR | 14 | 1.5 | 4 |
| HsKIF3A | K-2 | LRYANRA | KNIKNKARINEDPKDAL | LRQFQKE | 17 | 1 | 4 |
| HsKIF1A | K-3 | LRYADRT | KQIRCNAVINEDPNNKL I | IRELKDE | 17 | 0.5*/9.2** | 6 |
| HsKIF7 | K-4 | LNYASRA | QNIRNRATVNWRPE--- | AERPPEE | 14 | 0/nd | 本記事 |
| HsKIF11(Eg5, KSP) | K-5 | LEYAHRA | KNILNKPEVNQKLTKKAL | IKEYTEE | 18 | 0.3 | 8 |
*モノマー、**二量体、NL = ネックリンカー
Soppina ら6 によって K-3 ファミリーメンバー(例: KIF1A、KIF13A、KIF131B、KIF16B)の解析が行われ、コイルドコイル1サブドメイン(CC1)を NL 上に折り曲げると、processivity が非常に低下し、二量体化が阻害されることが示されました。K-3 ファミリーのメンバーは、一般的に processivity が低いとされていますが、細胞の末端部分に小胞を輸送する小胞モーターとしての機能に反するように思われます。Soppina ら6 は、NL-CC1 サブドメインとコイルドコイルを交換すると、processivity の高い二量体構造をとることから、モーターは、実際には in vivo でカーゴに結合して二量体化が起こると、processivity が高くなることを証明しました7。
また、Hesse ら8 によって、キネシンサブドメインの複雑な相互作用が報告されています。K-1 は、3つのサブドメイン[カバー、NL、L13(L5 アナログ)9,10 (図1)]を含むと考えられています。これらのサブドメインは、Pi 放出によって生じる力を NL に協調させることで、モータードメインの運動に寄与します。Hesse ら8 は、K-1 から K-5 ファミリーのメンバー間でこれらの3つのサブドメインをスワップすることで、「K-5 メンバーのドメインが K-1 に移行されると、K-5 メンバーに特徴的な低い弾力性を示す」といった、各モーター特異的な機能に関係する特徴をいくつか発見しています。
これらの研究から、キネシンサブドメインには互換性があり、本来のモーターの機能に関する情報を運んでいる、という新しい段階の知見が得られます。これらのサブドメインは、短いアミノ酸のストレッチで、モーター特異的な機能に大きく影響していると考えられます。また、これらの部位は、創薬および分子生物学のツールに関する潜在的な標的となります。
参考文献
1. Case R.B. et al. 2000. Role of the kinesin neck linker and catalytic core in microtubule-based motility. Curr. Biol.10, 157-160.
2. Hoeprich G.J. et al. 2014. Kinesin’s neck-linker determines its ability to navigate obstacles on the microtubule surface. Biophys. J. 106, 1691-1700.
3. Düselder A. et al. 2012. Neck-linker length dependence of processive kinesin-5 motility. J. Mol. Biol. 423, 159-168.
4. Shastry S. and Hancock W.O. 2010. Neck linker length determines the degree of processivity in kinesin-1 and kinesin-2 motors. Curr. Biol. 20, 939-943.
5. Shastry S. and Hancock W.O. 2011. Interhead tension determines processivity across diverse N-terminal kinesins.Proc. Natl. Acad. Sci. U.S.A. 108, 16253-16258.
6. Soppina V. et al. 2014. Dimerization of mammalian kinesin-3 motors results in superprocessive motion. Proc. Natl. Acad. Sci. U.S.A. 111, 5562-5567.
7. Tomishige M. et al. 2002. Conversion of UNC104/KIF1A kinesin into a processive motor after dimerization.Science. 297, 2263-2276.
8. Hesse W.R. et al. 2013. Modular aspects of kinesin force generation machinery. Biophys. J. 104, 1969-1978.
9. Khalil A.D. et al. 2008. Kinesin’s cover neck bundle folds forward to generate force. Proc. Natl. Acad. Sci. U.S.A.105, 19247-19252.
10. Hwang W. et al. 2008. Force generation in kinesin hinges on cover-neck bundle formation. Structure. 16, 62-71.
モータータンパク質
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
Myosin S1 Fragment (Cardiac), Bovine |
CYT | CS-MYS03 | 1*250 UG |
CYT社 CSMYS03A 1*250 を参照 |
| Myosin - smooth muscle S1 fragment, Chicken |
CYT | CS-MYS05 | 1*250 UG |
¥120,000 |
| Myosin, Rabbit |
CYT | CS-MYS04 | 1*250 UG |
¥120,000 |
| KIF7 motor domain, Human |
CYT | CS-KF51 | 1*100 UG |
¥150,000 |
| Cytoplasmic Dynein Motor Protein, Porcine |
CYT | CS-DN01 | 1*50 UG |
¥153,000 |
タンパク質・試薬
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
| Paclitaxel (Taxol), Plant |
CYT | TXD01 | 10*100 UL |
¥73,000 |
| Kinesin heavy chain motor domain protein (KIF5B), Human |
CYT | KR01-A | 2*25 UG |
¥150,000 |
| Kinesin heavy chain motor domain protein (KIF5B), Human |
CYT | KR01-XL | 1*1 MG |
お問い合わせ |
| CENP-E kinesin motor domain protein (KIF10), Human |
CYT | CP01-A | 2*25 UG |
¥150,000 |
| CENP-E kinesin motor domain protein (KIF10), Human |
CYT | CP01-XL | 1*1 MG |
お問い合わせ |
| Eg5 kinesin motor domain protein (KIF11), Human |
CYT | EG01-A | 2*25 UG |
¥150,000 |
| Eg5 kinesin motor domain protein (KIF11), Human |
CYT | EG01-B | 10*25 UG |
お問い合わせ |
| Eg5 kinesin motor domain protein (KIF11), Human |
CYT | EG01-XL | 1*1 MG |
お問い合わせ |
| Microtubules (Taxol Stabilized and Lyophilized), Porcine |
CYT | MT002-A | 4*500 UG |
¥153,000 |
| Microtubules (Taxol Stabilized and Lyophilized), Porcine |
CYT | MT002-XL | 1*10 MG |
お問い合わせ |
| Tubulin, Porcine |
CYT | T240-A | 1*1 MG |
¥45,000 |
| Tubulin, Porcine |
CYT | T240-B | 5*1 MG |
¥200,000 |
| Tubulin, Porcine |
CYT | T240-C | 20*1 MG |
お問い合わせ |
| Tubulin, Porcine |
CYT | T240-DX | 1*10 MG |
¥307,000 |
キット
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 |
|---|---|---|---|---|
| HTS kinesin ATPase Endpoint Assay Biochem Kit |
CYT | BK053 | 1 KIT [1000 assays] |
¥307,000 |
| Kinesin ELIPA kit |
CYT | BK060 | 1 KIT [96 assays] |
¥278,000 |
■ CYTOSKELETON NEWS バックナンバー
- 2020年10月号 紡錘体 - 可視化に向けた新規ツール
- 2020年8月号 細胞膜染色用蛍光プローブ
- 2020年4月号 生細胞におけるF-アクチンプローブ
- 2020年3月号 コロナウイルスと細胞骨格
- 2020年2月号 タウ(Tau)の将来性をMapping
- 2020年1月号 Rho GTPaseによる細胞遊走制御
- 2019年12月号 表現型プロファイリング:アクチンに焦点を当てたがん治療
- 2019年11月号 チューブリンの過剰グルタミル化、ミトコンドリア、神経変性
- 2019年9月号 細胞運動性を制御するために相互作用する膜張力とアクチン細胞骨格
- 2019年8月号 Rac1B、がん、およびRac1
- 2019年7月号 Rhoファミリー GTPases、神経可塑性、およびうつ状態
- 2019年6月号 アクチンメチオニン酸化: 動的制御の次の段階
- 2019年5月号 ミクログリアと神経変性疾患
- 2019年2月号 生細胞画像化に対するCNS疾患や障害
2018年
- 2018年12月号 アクチン細胞骨格とメカノトランスダクション(機械的シグナル伝達)
- 2018年11月号 軸索再生と細胞骨格
- 2018年10月号 ニューロンにおける微小管と極性
- 2018年8月号 Rab GTPase と 神経変性
- 2018年7月号 SUMO レスリング: バランスが全て
- 2018年6月号 なぜ K-Ras は発がん特異性を示すのか?
- 2018年5月号 治療標的としてのユビキチンプロテアソームシステム:チューブリンは関与するか?
- 2018年4月号 RhoファミリーGEFと樹状突起スパインの構造的可塑性
- 2018年3月号 βカテニンとTFC/LEF-1の翻訳後修飾による標準的なWntシグナル制御
- 2018年2月号 がん抑制遺伝子p53の翻訳後修飾による機能の調整
- 2018年1月号 自閉スペクトラム症におけるGEF Trioの役割
2017年
- 2017年12月号 プロフィリン: アクチン結合タンパク質の多機能な役割
- 2017年11月号 ミトコンドリアにおけるアセチル化:新たな考え方と治療への応用の可能性
- 2017年9月号 翻訳後修飾のアセチル化による微小管の安定化
- 2017年8月号 神経軸索におけるアクチンリングを基盤とした周期的膜骨格(PMS)
- 2017年7月号 E3ユビキチンリガーゼMdm2によるがん抑制遺伝子p53の翻訳後制御
- 2017年6月号 多能性幹細胞(PSC)での転写因子による翻訳後制御
- 2017年5月号 Arf6 GEF と癌細胞の浸潤・転移
- 2017年4月号 PTEN(Phosphatase and Tensin Homolog)による翻訳後制御
- 2017年3月号 Tau の翻訳後修飾: アルツハイマー病の治療標的
- 2017年2月号 樹状細胞の移動におけるアクチン結合タンパク質とF-アクチン
2016年
- 2016年11月/12月号 GEF を介した GTPase シグナル伝達の低分子阻害剤
- 2016年9月号 FtsZ タンパク質: 抗菌薬の新規ターゲット
- 2016年7月号 翻訳後修飾(PTM)は心臓病において細胞骨格タンパク質を調節する
- 2016年6月号 モータータンパク質キネシンと神経変性
- 2016年5月号 チロシンリン酸化は Rhoファミリー GTPase 活性を調節する
- 2016年4月号 Rac1と糖尿病: ポジティブな役割とネガティブな役割
- 2016年3月号 SUMO化: 細胞骨格タンパク質の機能を調節するレギュレーター
- 2016年1月/2月号 ビメンチン中間径フィラメント: リン酸化による調節
2015年
- 2015年8月号 タンパク質調節に不可欠な翻訳後修飾
- 2015年7月号 アクチン細胞骨格のライブセルイメージング
- 2015年6月号 有糸分裂に関わるタンパク質のSUMO化: 局在と機能
- 2015年5月号 Ras 癌の治療: 5つの有望なターゲット
- 2015年4月号 Ras 依存性の癌で注目される YAP1
- 2015年3月号 増刊号 統合失調症において遺伝子変異により誘導されるアクチン依存のシナプスの変化
- 2015年3月号 Ral GTPase を調節する翻訳後修飾
- 2015年1月/2月号 RhoA のリン酸化はシグナル伝達を調節する
- 2015年1月号 増刊号 微小管を不安定化する suprafenacine: 新規抗癌剤のリード化合物としての可能性
2014年
- 2014年12月号 増刊号 RhoA は心筋細胞におけるアクチン細胞骨格の再構成とグルコース取り込みを仲介する
- 2014年11月号 増刊号 樹状突起の形態形成: ドーパミンD1受容体 および Rho ファミリー GTPase による制御
- 2014年11月/12月号 GTPase 活性化アッセイ: アイソフォームの検出
- 2014年10月号 アルギニンの正電荷を消失させるシトルリン化
- 2014年9月号 キネシンサブドメインの探索
- 2014年9月号 増刊号 アクチン結合タンパク質コフィリンの S-ニトロシル化: 細胞移動に対する影響
- 2014年8月号 増刊号 原発性硬化性胆管炎における N-Ras 発現および活性
- 2014年8月号 SUMO化: 細胞骨格タンパク質を標的とした翻訳後修飾
- 2014年7月号 Sos/K-Ras 結合を介して Ras シグナル伝達を制御する新しい低分子阻害剤
- 2014年6月号 増刊号 頭頸部扁平上皮癌における microRNA-138 による RhoC のダウンレギュレーション
- 2014年6月号 Rho GTPase と活性酸素種: クロストークとフィードバック
- 2014年5月号 ミオシンのアセチル化はサルコメアの構造と機能を調節する
- 2014年4月号 リジンのアセチル化 - 多様な細胞プロセスの制御因子
- 2014年3月号 インテグリンを介したβ-アクチンの酸化還元制御: PDIの出現
- 2014年1/2月号 ダイニン: 一つのモーターが関わる複数の神経変性疾患
2013年
- 2013年11/12月号 ダイニン:チームとして強力に作用するモータータンパク質
- 2013年10月号 神経変性:Rhes、SUMO化、ハンチントン病
- 2013年9月号 モノユビキチン化:タンパク質調節のダイナミックなタグ
- 2013年8月号 Ras及びRhoのプレニル化による翻訳後修飾:癌創薬における役割
- 2013年7月号 アクチンが引き起こす膜突起による浸潤:コルタクチン
- 2013年6月号 アクチン修飾と細胞骨格
- 2013年5月号 微小管内部の実体
- 2013年4月号 神経変性におけるTauの多面性
- 2013年3月号 蛍光フィブロネクチンタンパク質を用いた特発性肺線維症の創薬
- 2013年1/2月号 樹状突起棘:発生におけるArf6の役割
2012年
- 2012年11/12月号 ミオシンの小分子モジュレーター
- 2012年10月号 Rhoファミリーパスウェイのユビキチン化と制御
- 2012年9月号 神経変性におけるRac1 GTPaseの機能
- 2012年8月号 上皮間葉転換(EMT)とRhoファミリー低分子量G-タンパク質の関与
- 2012年7月号 チューブリンの多重修飾:グルタミル化とグリシル化
- 2012年6月号 細胞接着のフィブロネクチン制御と原線維形成
- 2012年5月号 アクチン酸化サイクルの機能
- 2012年4月号 トラフィッキング:ArfとCdc42/Racの結合
- 2012年3月号 G-LISAを用いた心臓研究: 糖尿病性心筋症におけるRho経路に関する研究
- 2012年1月/2月号 FtsZ: 新たな抗生剤の標的となるチュ−ブリンホモログ
商品は「研究用試薬」です。人や動物の医療用・臨床診断用・食品用としては使用しないように、十分ご注意ください。
※ 表示価格について














