






















 このページを印刷する
このページを印刷する
記事ID : 17803
Arf6 GEF と癌細胞の浸潤・転移 CYTOSKELETON NEWS 2017年5月号
Arf6 GEF と癌細胞の浸潤・転移
はじめに
低分子量Gタンパク質 Arf6(ADP-ribosylation factor 6)は Ras スーパーファミリーのうち Arf サブファミリーに属しています。Arf GTPase は3つのクラスに分類されますが、Arf6 は唯一クラスIIIに分類され、細胞膜やエンドソームに局在しアクチン細胞骨格のダイナミクスに基づく様々な細胞運動(エンドサイトーシス、エキソサイトーシス、膜局在タンパク質の輸送や再循環、細胞膜のラフリングなど)を制御しています。これらの細胞機能は生理学的/病理学的な細胞運動および細胞内輸送の基盤となっています。Arf6 は、GDPと結合した不活性状態とGTPと結合した活性状態との間を行き来し、上述の細胞プロセスにおける分子スイッチとして機能しています。Arf6 は、GEF(guanine exchange factors)によりGDPがGTPへと変換されると活性化され、GAP(GTPase activating proteins)によりGTPが加水分解されると不活性化されます1-3。 本稿では、癌細胞の浸潤と転移における Arf6 と Arf6 GEF の役割に注目します。
Arf6 GEFと癌
癌細胞の浸潤と転移にはアクチン細胞骨格の動的な変化が関与しています。最近の研究では、Arf6が癌細胞の浸潤や転移だけでなく腫瘍の血管新生や増殖においても重要な役割を担っていることが報告されています4-8。さらに BRAG2/GEP100 や cytohesin3/Grp1、EFA6 のような Arf6 GEF は癌の進行にも関与しており5,7,8、Arf6 の活性やサイクルを阻害することで癌の進行を抑制できます6,7,9。
Arf GEF は14種類が報告されています。そのうち8種類が Arf6 を標的にしており、さらにそのうち5種類は Arf6 特異的です。Arf6 GEF は、さまざまなリガンドにより活性化される細胞表面受容体と同様に、PH(pleckstrin homology)ドメインと PI3K(phosphatidylinositol 3-kinase)により生成される phosphoinositide とが直接結合することで活性化され、細胞膜にリクルートされます。GEFの触媒作用を担う Sec7 ドメインは、膜に局在する Arf6 を活性化します。BRAG2/GEP100 や EFA6A-D は Arf6 のみを活性化しますが、cytohesin1 や cytohesin2/ARNO、cytohesin3/Grp1 は Arf6 および他の Arf を標的としています1-3。
GEP100 は Arf6 GEF の中でも解析が進んでおり、乳癌やメラノーマ、肺腺癌の浸潤を促進することがわかっています5,6,10,11。 EGF(epidermal growth factor)刺激に応じて、GEP100 はリガンドの結合した EGFR(EGF receptor)と直接結合し、細胞膜の Arf6 を活性化します5(図1)。次にGTP結合型 Arf6 が下流のエフェクター分子である AMAP1 をリクルートし活性化させます。これにより浸潤突起が形成され、癌細胞浸潤の準備が整い始めます10。活性化された Her2(human epidermal growth factor 2)もまた GEP100 を介した Arf6 活性化を刺激し、癌細胞浸潤を正に制御します11。VEGF(vascular endothelial growth factor)も同様に GEP100 を介して Arf6 を活性化し、内皮細胞における血管新生を促進します12。GEP100 を介した Arf6 の活性化は、E-cadherin を介した細胞間接着を分解させることでも癌細胞の浸潤や転移を促進します10(図1)。また、メラノーマ細胞ではWNT5A(Wnt family member 5A)が GEP100 を介して Arf6 を活性化し、これにより β-catenin/cadherin 複合体が分解されることで癌細胞の浸潤や転移が促進されることが報告されています6(図1)。これらの作用は、GEP100 や Arf GEF の cytohesin ファミリーの低分子阻害剤 SecinH3 により阻害されます6。
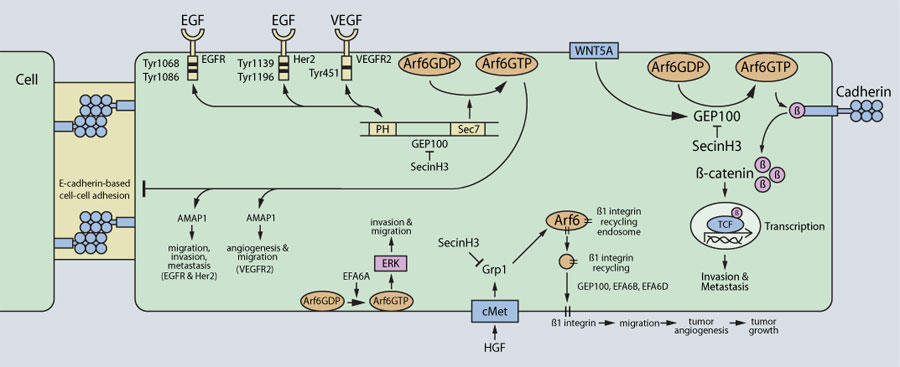
図1 Arf6 GEFのシグナル伝達経路と癌細胞の浸潤、転移、腫瘍血管新生
Arf6 GEF の EFA6 ファミリーでは、EFA6 GEF アイソフォームや癌種に応じて正負の制御が異なっています。 例えば、EFA6A はグリオーマ細胞の浸潤を亢進しますが13、EFA6B は乳癌の浸潤を抑制します14。EFA6A を介した作用はERK(extracellar signal-regulated kinase)シグナル伝達経路のカスケードを用いており13、Arf6 刺激によるメラノーマ細胞の浸潤にはERKシグナル伝達が必要であるという報告もあります15。複雑なことに、EFA6A, B, C アイソフォームは癌細胞の浸潤や転移、薬剤抵抗性を亢進しています8。
Grp1 を介した Arf6 の活性化は、b1インテグリンのエンドソームから細胞膜へのリサイクリングを増加させることで、HGF(Hepatocyte growth factor)依存的な腫瘍の血管新生を制御します7(図1)。b1インテグリンの細胞膜へのリサイクリングは、形成された新生血管の成熟化を促す癌細胞の固定に関与します。GEP100 と同様に、SecinH3 によりこれらは抑制されます7。Grp1 は乳癌細胞の遊走もコントロールすることが報告されています16。
臨床的な意義:Arf6の低分子阻害剤
GEP100 や cytohesin の阻害は、上述の通り Arf GEF の低分子阻害剤 SecinH3 を用いることで可能です。しかしながら動物試験では、SecinH3 はグリオーマ細胞の転移やメラノーマ、肺腺癌の血管新生を抑制できていますが6,7、肝臓のインスリン抵抗性が上昇することも報告されています17。 別の低分子阻害剤である PIT-1 は、細胞の生存や増殖に関わる Akt の制御因子でありPI3Kの生成物であるPIP3(phosphatidylinositol-3,4,5-triphosphate)を阻害します。 PIT-1は、ARNOや Grp1 のPHドメインとPIP3の結合を妨害することでラメリポディア形成や乳癌細胞の遊走を阻害し1,2,16、メラノーマにおける血管新生や転移を抑制します。癌細胞の浸潤や転移においてARNOと Grp1 のどちらが重要か、PIT-1 による抑制効果がどちらに、もしくは両方によるものなのかなどについてはさらなる研究が必要です。
まとめ
Arf6 GTPase やGEFの最近の研究は、これらのタンパク質が癌細胞の浸潤や転移、腫瘍血管新生において明確な役割を持っていることを示しており、実際に幾つかの癌種においては特定のGEFが関与していることも判明しています。これらの知見は、Arf6 や Arf6 GEF が選択的かつ新しい抗癌剤の標的となり得ることを示唆しています。 これらの研究をサポートするために、Cytoskeleton社ではこれまでの Ras スーパーファミリーの研究キットに加え Arf6 activation assay キットをご提供しています。この activation assay は、エフェクタービーズを用いたプルダウンアッセイとELISA法に基づくG-LISAアッセイの二種類のフォーマットが利用できます。また、GEF/GAP exchange アッセイキットや、生細胞/固定細胞の F-actin の変化をモニターできる試薬も提供しています。
参考文献
- Yamauchi Y. et al. 2017. Machineries regulating the activity of the small GTPase Arf6 in cancer cells are potential targets for developing innovative anti-cancer drugs. Adv. Biol. Regul. 63, 115-121.
- Hongu T. and Kanaho Y. 2014. Activation machinery of the small GTPase Arf6. Adv Biol. Regul. 54, 59-66.
- Donaldson J.G. et al. 2011. ARF family G proteins and their regulators: roles in membrane transport, development and disease. Nat. Rev. Mol. Cell Biol. 12, 362-375.
- Hashimoto S. et al. 2004. Requirement for Arf6 in breast cancer invasive activities. Proc. Natl. Acad. Sci. U.S.A.101, 6647-6652.
- Morishige M. et al. 2008. GEP100 links epidermal growth factor receptor signalling to Arf6 activation to induce breast cancer invasion. Nat. Cell Biol. 10, 85-92.
- Grossmann A.H. et al. 2013. The small GTPase Arf6 stimulates beta-catenin transcriptional activity during WNT5A-mediated melanoma invasion and metastasis. Sci. Signal. 6, ra14.
- Hongu T. et al. 2015. Arf6 regulates tumour angiogenesis and growth through HGF-induced endothelial b1 integrin recycling. Nat. Commun. 6, 7925.
- Hashimoto S. et al. 2016. Lysophosphatidic acid activates Arf6 to promote the mesenchymal malignancy of renal cancer. Nat. Commun. 7, 10656.
- Yoo J.H. et al. 2016. ARF6 is an actionable node that orchestrates oncogenic GNAQ signaling in uveal melanoma. Cancer Cell. 29, 1-16.
- Sabe H. et al. 2009. The EGFR-GEP100-Arf6-AMAP1 signaling pathway specific to breast cancer invasion and metastasis. Traffic. 10, 982-993.
- Menju T. et al. 2011. Engagement of overexpressed Her2 with GEP100 induces autonomous invasive activities and provides a biomarker for metastases of lung adenocarcinoma. PLoS One. 6, e25301.
- Hashimoto A. et al. 2011. GEP100-Arf6-AMAP1-Cortactin pathway frequently used in cancer invasion is activated by VEGFR2 to promote angiogenesis. PLoS ONE. 6, e23359.
- Li M. et al. 2006. EFA6A enhances glioma cell invasion through ADP ribosylation factor 6/extracellular signal-regulated kinas signaling. Cancer Res. 66, 1583-1590.
- Zangari J. et al. 2014. EFA6B antagonizes breast cancer. Cancer Res. 74, 5493-5506.
- Tague S.E. et al. 2004. ADP-ribosylation factor 6 regulates tumor cell invasion through the activation of the MEK/ERK signaling pathway. Proc. Natl. Acad. Sci. U.S.A. 101, 9671-9676.
- Miao B. et al. 2012. Inhibition of cell migration by PITENINs: the role of ARF6. Oncogene. 31, 4317-4332.
- Hafner M. et al. 2006. Inhibition of cytohesins by SecinH3 leads to hepatic insulin resistance. Nature. 444,941-944.
Arf 活性アッセイキット
[商品詳細]
- Arf1 / Arf6 活性アッセイキット
Arf1とArf6の活性を2種類(G-LISA、プルダウン)の方法で測定
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 | 在庫 |
|---|---|---|---|---|---|
Arf1 Pull-down Activation Assay Biochem Kit (bead pull-down format) |
CYT | BK032-S | 20 ASSAY |
¥170,000 | なし |
| Arf6 Pull-down Activation Assay Biochem Kit (bead pull-down format) |
CYT | BK033-S | 20 ASSAY |
¥170,000 | なし |
| Arf1 G-LISA Activation Assay Kit |
CYT | BK132 | 96 ASSAY |
販売終了 | 販売終了 |
| Arf6 G-LISA Activation Assay Kit |
CYT | BK133 | 96 ASSAY |
販売終了 | 販売終了 |
F-アクチン視覚化試薬
[商品詳細]
- Spirochrome プローブ(SiR-Actin/SiR-Tubulin/SiR-DNA)
- Spirochrome プローブ(SiR700-Actin / SiR700-Tubulin / SiR700-DNA)
細胞骨格(アクチン・チューブリン)やDNAを超解像度で蛍光観察 - Acti-Stain™ 蛍光標識ファロイジン
生細胞/固定細胞の細胞骨格研究に有用
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 | 在庫 |
|---|---|---|---|---|---|
| SiR-Actin Kit |
CYT | CY-SC001 | 1 KIT [50-300 slides] |
¥182,000 | なし |
| SiR700-Actin Kit |
CYT | CY-SC013 | 1 KIT [35-200 slides] |
¥182,000 | なし |
| Acti-stain 488 phalloidin, Plant |
CYT | PHDG1-A | 1*500 UL [300 slides] |
¥79,000 | あり |
| Acti-stain 555 phalloidin, Mushroom |
CYT | PHDH1-A | 1*500 UL [300 slides] |
¥79,000 | あり |
| Rhodamine Phalloidin, Rhodamine Isothiocyanate |
CYT | PHDR1 | 1*500 UL [300 slides] |
¥79,000 | あり |
その他の生細胞イメージングプローブ
[商品詳細]
- Spirochrome プローブキット(SiR-Lysosome / SiR700-Lysosome)
生細胞中のリソソームを far-red(遠赤色光)でイメージング
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 | 在庫 |
|---|---|---|---|---|---|
| SiR-DNA Kit |
CYT | CY-SC007 | 1 KIT [50-300 slides] |
¥120,000 | あり |
| SiR-Lysosome Kit |
CYT | CY-SC012 | 1 KIT [50-300 slides] |
¥131,000 | なし |
■ CYTOSKELETON NEWS バックナンバー
- 2020年10月号 紡錘体 - 可視化に向けた新規ツール
- 2020年8月号 細胞膜染色用蛍光プローブ
- 2020年4月号 生細胞におけるF-アクチンプローブ
- 2020年3月号 コロナウイルスと細胞骨格
- 2020年2月号 タウ(Tau)の将来性をMapping
- 2020年1月号 Rho GTPaseによる細胞遊走制御
- 2019年12月号 表現型プロファイリング:アクチンに焦点を当てたがん治療
- 2019年11月号 チューブリンの過剰グルタミル化、ミトコンドリア、神経変性
- 2019年9月号 細胞運動性を制御するために相互作用する膜張力とアクチン細胞骨格
- 2019年8月号 Rac1B、がん、およびRac1
- 2019年7月号 Rhoファミリー GTPases、神経可塑性、およびうつ状態
- 2019年6月号 アクチンメチオニン酸化: 動的制御の次の段階
- 2019年5月号 ミクログリアと神経変性疾患
- 2019年2月号 生細胞画像化に対するCNS疾患や障害
2018年
- 2018年12月号 アクチン細胞骨格とメカノトランスダクション(機械的シグナル伝達)
- 2018年11月号 軸索再生と細胞骨格
- 2018年10月号 ニューロンにおける微小管と極性
- 2018年8月号 Rab GTPase と 神経変性
- 2018年7月号 SUMO レスリング: バランスが全て
- 2018年6月号 なぜ K-Ras は発がん特異性を示すのか?
- 2018年5月号 治療標的としてのユビキチンプロテアソームシステム:チューブリンは関与するか?
- 2018年4月号 RhoファミリーGEFと樹状突起スパインの構造的可塑性
- 2018年3月号 βカテニンとTFC/LEF-1の翻訳後修飾による標準的なWntシグナル制御
- 2018年2月号 がん抑制遺伝子p53の翻訳後修飾による機能の調整
- 2018年1月号 自閉スペクトラム症におけるGEF Trioの役割
2017年
- 2017年12月号 プロフィリン: アクチン結合タンパク質の多機能な役割
- 2017年11月号 ミトコンドリアにおけるアセチル化:新たな考え方と治療への応用の可能性
- 2017年9月号 翻訳後修飾のアセチル化による微小管の安定化
- 2017年8月号 神経軸索におけるアクチンリングを基盤とした周期的膜骨格(PMS)
- 2017年7月号 E3ユビキチンリガーゼMdm2によるがん抑制遺伝子p53の翻訳後制御
- 2017年6月号 多能性幹細胞(PSC)での転写因子による翻訳後制御
- 2017年5月号 Arf6 GEF と癌細胞の浸潤・転移
- 2017年4月号 PTEN(Phosphatase and Tensin Homolog)による翻訳後制御
- 2017年3月号 Tau の翻訳後修飾: アルツハイマー病の治療標的
- 2017年2月号 樹状細胞の移動におけるアクチン結合タンパク質とF-アクチン
2016年
- 2016年11月/12月号 GEF を介した GTPase シグナル伝達の低分子阻害剤
- 2016年9月号 FtsZ タンパク質: 抗菌薬の新規ターゲット
- 2016年7月号 翻訳後修飾(PTM)は心臓病において細胞骨格タンパク質を調節する
- 2016年6月号 モータータンパク質キネシンと神経変性
- 2016年5月号 チロシンリン酸化は Rhoファミリー GTPase 活性を調節する
- 2016年4月号 Rac1と糖尿病: ポジティブな役割とネガティブな役割
- 2016年3月号 SUMO化: 細胞骨格タンパク質の機能を調節するレギュレーター
- 2016年1月/2月号 ビメンチン中間径フィラメント: リン酸化による調節
2015年
- 2015年8月号 タンパク質調節に不可欠な翻訳後修飾
- 2015年7月号 アクチン細胞骨格のライブセルイメージング
- 2015年6月号 有糸分裂に関わるタンパク質のSUMO化: 局在と機能
- 2015年5月号 Ras 癌の治療: 5つの有望なターゲット
- 2015年4月号 Ras 依存性の癌で注目される YAP1
- 2015年3月号 増刊号 統合失調症において遺伝子変異により誘導されるアクチン依存のシナプスの変化
- 2015年3月号 Ral GTPase を調節する翻訳後修飾
- 2015年1月/2月号 RhoA のリン酸化はシグナル伝達を調節する
- 2015年1月号 増刊号 微小管を不安定化する suprafenacine: 新規抗癌剤のリード化合物としての可能性
2014年
- 2014年12月号 増刊号 RhoA は心筋細胞におけるアクチン細胞骨格の再構成とグルコース取り込みを仲介する
- 2014年11月号 増刊号 樹状突起の形態形成: ドーパミンD1受容体 および Rho ファミリー GTPase による制御
- 2014年11月/12月号 GTPase 活性化アッセイ: アイソフォームの検出
- 2014年10月号 アルギニンの正電荷を消失させるシトルリン化
- 2014年9月号 キネシンサブドメインの探索
- 2014年9月号 増刊号 アクチン結合タンパク質コフィリンの S-ニトロシル化: 細胞移動に対する影響
- 2014年8月号 増刊号 原発性硬化性胆管炎における N-Ras 発現および活性
- 2014年8月号 SUMO化: 細胞骨格タンパク質を標的とした翻訳後修飾
- 2014年7月号 Sos/K-Ras 結合を介して Ras シグナル伝達を制御する新しい低分子阻害剤
- 2014年6月号 増刊号 頭頸部扁平上皮癌における microRNA-138 による RhoC のダウンレギュレーション
- 2014年6月号 Rho GTPase と活性酸素種: クロストークとフィードバック
- 2014年5月号 ミオシンのアセチル化はサルコメアの構造と機能を調節する
- 2014年4月号 リジンのアセチル化 - 多様な細胞プロセスの制御因子
- 2014年3月号 インテグリンを介したβ-アクチンの酸化還元制御: PDIの出現
- 2014年1/2月号 ダイニン: 一つのモーターが関わる複数の神経変性疾患
2013年
- 2013年11/12月号 ダイニン:チームとして強力に作用するモータータンパク質
- 2013年10月号 神経変性:Rhes、SUMO化、ハンチントン病
- 2013年9月号 モノユビキチン化:タンパク質調節のダイナミックなタグ
- 2013年8月号 Ras及びRhoのプレニル化による翻訳後修飾:癌創薬における役割
- 2013年7月号 アクチンが引き起こす膜突起による浸潤:コルタクチン
- 2013年6月号 アクチン修飾と細胞骨格
- 2013年5月号 微小管内部の実体
- 2013年4月号 神経変性におけるTauの多面性
- 2013年3月号 蛍光フィブロネクチンタンパク質を用いた特発性肺線維症の創薬
- 2013年1/2月号 樹状突起棘:発生におけるArf6の役割
2012年
- 2012年11/12月号 ミオシンの小分子モジュレーター
- 2012年10月号 Rhoファミリーパスウェイのユビキチン化と制御
- 2012年9月号 神経変性におけるRac1 GTPaseの機能
- 2012年8月号 上皮間葉転換(EMT)とRhoファミリー低分子量G-タンパク質の関与
- 2012年7月号 チューブリンの多重修飾:グルタミル化とグリシル化
- 2012年6月号 細胞接着のフィブロネクチン制御と原線維形成
- 2012年5月号 アクチン酸化サイクルの機能
- 2012年4月号 トラフィッキング:ArfとCdc42/Racの結合
- 2012年3月号 G-LISAを用いた心臓研究: 糖尿病性心筋症におけるRho経路に関する研究
- 2012年1月/2月号 FtsZ: 新たな抗生剤の標的となるチュ−ブリンホモログ
商品は「研究用試薬」です。人や動物の医療用・臨床診断用・食品用としては使用しないように、十分ご注意ください。
※ 表示価格について














