






















 このページを印刷する
このページを印刷する
記事ID : 9247
アクチン酸化サイクルの機能 CYTOSKELETON NEWS 2012年5月号
アクチン酸化サイクルの機能
in vivoにおけるアクチン細胞骨格の機能と編成が高度に動的なため、細胞内外のシグナルに速やかに応答することができます。アクチンの再編成は走化性、細胞の増殖や分裂、軸索伸長、貪食といった細胞の生理的なプロセスのほとんどで必須です。これら事象を複雑化するため、多様なアクチン制御因子が要求されます。よく知られた制御因子として、脂質キナーゼ/ホスファターゼ、小分子Gタンパク質であるRacやRhoファミリーといった転写因子と同様に、プロフィリン、ゲルソリン、コフィリンのようなアクチン結合タンパク質が挙げられます(1)。最近、主に過酸化水素(H2O2)が関与するアクチンの酸化サイクルが強力な制御機構であることが明らかになりました。H2O2のような活性酸素種は、NADPH酸化酵素やリポキシゲナーゼを活性化する多様な細胞外シグナルイベントにより産生されます。in vivoで、アクチンは酸化還元に感受性のある2か所の主要なシステイン残基、β-アクチンではCys272とCys374を反応中心として、還元型(actR)と酸化型(actO)の間を平衡します(2, 3、図1)。Cys272はH2O2存在下でスルフェン酸(Cys-SOH)、スルフィン酸(Cys-SO2H)、またはスルホン酸(Cys-SO3H)に酸化されます。Cys374は同様に酸化されるか、ジスルフィド結合を形成してアクチン二量体となります(4)。これら改変はチオレドキシン/チオレドキシン還元酵素/NADPH(TRX)やグルタチオン/グルタチオン還元酵素/NADPH(GLT)が関与する還元性の細胞経路により還元されるため、可逆的です。
興味深いことに、これら2つの酸化型システインが酸化型アクチンに重要な生理作用を与えています。例えば、actOはプロフィリンとの親和性が約10倍低下し(Cys374の酸化)、in vitroでのactO形成は還元剤の添加により可逆性を示す既成フィラメントの脱重合を引き起こします(3)。仮説上、これら2つのactOの特性により、局所的な酸化環境にて短く動的なフィラメントの形成が促進されます。加えて、in vitroでのH2O2によるCys374の酸化はアクチン二量体の形成を促進し、凝集中心を作成して分枝フィラメントを形成するためフィラメントに取り込まれます(4)。Arp2/3タンパク質はこれら構造の安定化と娘フィラメントの形成に関与すると考えられます(3)。最後に、Cys272の酸化によるスルフェン酸の形成はアポトーシスによる細胞死を決定づける酸化還元のセンサーとなっています(5)。
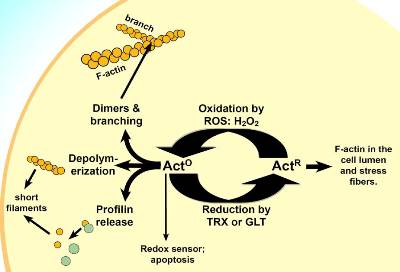
図1 in vivoでのactOの潜在的な機能
ダイマーと短いフィラメントの形成能を有するactOと、in vivoにおけるアクチンの酸化型と還元型との循環。actOは酸化状態の場合にアポトーシスを誘導する、細胞内酸化還元電位のセンサーでもある。TRX、チオレドキシン/チオレドキシン還元酵素/NADPH; GLT、グルタチオン/グルタチオン還元酵素/NADPH
アクチン酸化サイクルにおいて、他に興味深い側面としては、低分子Gタンパク質であるRacがアクチン制御において、よく研究されたRac/エフェクター関連の成長円錐動態のコントロールに加え、他の役割も有する可能性が挙げられます(6)。RacはH2O2濃度を増幅するシグナルカスケードに関与します(7, 8)。H2O2による脂質やタンパク質チロシンのキナーゼホスファターゼ(例、PTEN)のダウンレギュレーションは、PIP2とPIP3の量を増加させます(9)。これらホスホイノシチドがRacを活性化するグアニン交換因子を標的とし、NADPH酸化酵素を活性化するので(12, 13)、その結果としてH2O2の産生が増加します(9)。そして時間と空間に関連して細胞表面で増幅されたH2O2のプールによりアクチンが酸化され、細胞表面のアクチン動態が制御されます。このトレッドミル用の反応によりアクチンが細胞内腔へ移行した後、actOはTRXやGLT還元系によりactRへと変換されます。
重要なことに、ヒトの疾患におけるactOの機能に関する2つの興味深いモデルが登場したため、アクチン酸化の研究は臨床的な意味があります。in vitroの炎症性腸疾患モデルにおいて、actOは大腸癌由来Caco-2細胞の腸管障壁の完全性を損傷するにあたり重要な役割を果たします(14)。また、虚血時、actOは心臓細胞のアポトーシスと細胞死に寄与します(15)。
参考文献
1. Sundaresan M, Yu ZX, Ferrans VJ, Irani K, and Finkel T. 1995. Requirement for generation of H2O2 for platelet-derived growth factor signal transduction. Science. 270, 296-299.
2. Fiaschi T, Cozzi G, Raugei G, Formigli L, Ramponi G, and Chiarugi P. 2006. Redox regulation of beta-actin during integrin-mediated cell adhesion. J. Biol. Chem. 281, 22983-22991.
3. Lassing I, Schmitzberger F, Bjornstedt M, Holmgren A, Nordlund P, Schutt CE, and Lindberg U. 2007. Molecular and structural basis for redox regulation of β-actin. J. Mol. Biol. 370, 331-348.
4. Tang JX, Janmey PA, Stossel TP, and Ito T. 1999. Thiol oxidation of actin produces dimers that enhance the elasticity of the F-actin network. Biophys. J. 76, 2208-2215.
5. Farah ME and Amberg DC. 2007. Conserved actin cysteine residues are oxidative stress sensors that can regulate cell death in yeast. Mol. Biol. Cell. 18, 1359-1365.
6. Hall A. 1998. Rho GTPases and the actin cytoskeleton. Science. 279, 509-514.
7. Moldovan L, Irani K, Moldovan NI, Finkel T, and Goldschmidt-Clermont PJ. 1999. The actin cytoskeleton reorganization induced by Rac1 requires the production of
superoxide. Antioxid. Redox Signal. 1, 29-43.
8. Rhee SG, Bae YS, Lee SR, and Kwon J. 2000. Hydrogen peroxide: a key messenger that modulates protein phosphorylation through cysteine oxidation. Sci. STKE. 2000, PE1.
9. Rhee SG, Kang SW, Jeong W, Chang TS, Yang KS, and Woo HA. 2005. Intracellular messenger function of hydrogen peroxide and its regulation by peroxiredoxins. Curr. Opin. Cell Biol. 17, 183-189.
10. Hawkins PT, Eguinoa A, Qiu R-G, Stokoe D, Cooke FT, Walters R, Wennstrom S, Claesson-Welsh L, Evans T, Symons M, and Stephens L. 1995. PDGF stimulates an increase in GTP-Rac via activation of phosphoinositide 3-kinase. Curr. Biol. 5, 393-403.
11. Han J, Luby-Phelps K, Das B, Shu X, Xia Y, Mosteller RD, Krishna UM, Falck JR, White MA, and Broek D. 1998. Role of substrates and products of PI3-kinase in regulating activation of Rac-related guanosine triphosphatases by Vav. Science. 279, 558-560.
12. Abo A, Pick E, Hall A, Totty N, Teahan CG, and Segal AW. 1991. Activation of the NADPH oxidase involves the small GTP-binding protein p21rac1. Nature. 353, 668-670.
13. Moldovan L, Mythreye K, Goldschmidt-Clermont PJ, and Satterwhite LL. 2006. Reactive oxygen species in vascular endothelial cell motility. Roles of NAD(P)H
oxidase and Rac1. Cardiovasc. Res. 71, 236-246.
14. Banan A, Zhang Y, Losurdo J, and Keshavarzian A. 2012. Carbonylation and disassembly of the F-actin cytoskeleton in oxidant induced barrier dysfunction and its prevention by epidermal growth factor and transforming growth factor a in a human colonic cell line. Gut. 46, 830-837.
15. Canton M, Neverova I, Menabo R, Van Eyk J, and Di Lisa F. 2004. Evidence of myofibrillar protein oxidation induced by postischemic reperfusion in isolated rat
hearts. Am. J. Physiol. Heart Circ. Physiol. 286, H870-H877.
アクチンBiochemキット™
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 | 在庫 |
|---|---|---|---|---|---|
Actin Polymerization Biochem Kit (fluorescence format), Rabbit |
CYT | BK003 | 1 KIT [30-100 assays] |
¥304,000 | なし |
| G-Actin/F-actin In Vivo Assay Biochem Kit, Mouse |
CYT | BK037 | 1 KIT [30-100 assays] |
¥264,000 | なし |
| Actin Binding Protein Spin-Down Assay Biochem Kit, Rabbit |
CYT | BK001 | 1 KIT [30-100 assays] |
¥268,000 | なし |
| Actin Binding Protein Spin-Down Assay Biochem Kit, Human |
CYT | BK013 | 1 KIT [30-100 assays] |
¥272,000 | なし |
| F-actin Visualization Biochem Kit (fluorescence format) |
CYT | BK005 | 1 KIT [300 slides] |
¥141,000 | なし |
Rac1活性化アッセイ&活性化因子
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 | 在庫 |
|---|---|---|---|---|---|
| Rac1 G-LISA(R) Activation Assay Kit |
CYT | BK128 | 1 KIT [96 assays] |
CYT社 BK128L 1 を参照 |
なし |
| Rho/Rac/Cdc42 Activator I |
CYT | CN04-A | 3*20 UG |
¥76,000 | なし |
| Rho/Rac/Cdc42 Activator I |
CYT | CN04-B | 9*20 UG |
¥211,000 | なし |
ファロイジン
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 | 在庫 |
|---|---|---|---|---|---|
| Acti-stain 488 phalloidin, Plant |
CYT | PHDG1-A | 1*500 UL [300 slides] |
¥79,000 | あり |
| Rhodamine Phalloidin, Rhodamine Isothiocyanate |
CYT | PHDR1 | 1*500 UL [300 slides] |
¥79,000 | あり |
| Acti-stain 555 phalloidin, Mushroom |
CYT | PHDH1-A | 1*500 UL [300 slides] |
¥79,000 | あり |
| Acti-stain 670 phalloidin, Plant |
CYT | PHDN1-A | 1*500 UL [300 slides] |
¥79,000 | あり |
アクチン結合タンパク質
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 | 在庫 |
|---|---|---|---|---|---|
| Alpha-actinin, Rabbit, Unlabeled |
CYT | AT01-A | 2*50 UG |
¥110,000 | なし |
| Alpha-actinin, Rabbit, Unlabeled |
CYT | AT01-C | 10*50 UG |
¥368,000 | なし |
| Cofilin 1, Human, Unlabeled |
CYT | CF01-A | 1*100 UG |
¥91,000 | なし |
| Cofilin 1, Human, Unlabeled |
CYT | CF01-C | 4*100 UG |
¥288,000 | なし |
| Gelsolin, Human |
CYT | HPG6-A | 4*20 UG |
¥123,000 | なし |
| Gelsolin, Human |
CYT | HPG6-B | 20*20 UG |
¥399,000 | なし |
| Myosin, Bovine |
CYT | MY03-A | 5*1 MG |
¥116,000 | なし |
| Myosin, Bovine |
CYT | MY03-B | 20*1 MG |
¥335,000 | なし |
| Heavy meromyosin, Rabbit, Unlabeled |
CYT | MH01-A | 4*50 UG |
¥102,000 | なし |
| Myosin II, Rabbit, Unlabeled |
CYT | MY02-A | 5*1 MG |
¥116,000 | なし |
| Myosin II, Rabbit, Unlabeled |
CYT | MY02-B | 20*1 MG |
¥335,000 | なし |
| Profilin 1, Human, Unlabeled |
CYT | PR01-A | 1*50 UG |
販売終了 | 販売終了 |
| WASP, Human |
CYT | VCG03-A | 1*500 UG |
¥177,000 | なし |
抗体
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 | 在庫 |
|---|---|---|---|---|---|
| Anti Actin w/positive control, Human (Rabbit) Unlabeled |
CYT | AAN01-A | 1*100 UG |
販売終了 | 販売終了 |
| Anti Actin w/positive control, Human (Rabbit) Unlabeled |
CYT | AAN01-B | 3*100 UG |
販売終了 | 販売終了 |
| Anti Cofilin w/positive control, Human (Rabbit) Unlabeled |
CYT | ACFL02-A | 1*50 UG |
販売終了 | 販売終了 |
| Anti Cofilin w/positive control, Human (Rabbit) Unlabeled |
CYT | ACFL02-B | 3*50 UG |
販売終了 | 販売終了 |
| Anti Profilin w/positive control, Human (Rabbit) Unlabeled |
CYT | APUF01-A | 1*50 UG |
販売終了 | 販売終了 |
■ CYTOSKELETON NEWS バックナンバー
- 2020年10月号 紡錘体 - 可視化に向けた新規ツール
- 2020年8月号 細胞膜染色用蛍光プローブ
- 2020年4月号 生細胞におけるF-アクチンプローブ
- 2020年3月号 コロナウイルスと細胞骨格
- 2020年2月号 タウ(Tau)の将来性をMapping
- 2020年1月号 Rho GTPaseによる細胞遊走制御
- 2019年12月号 表現型プロファイリング:アクチンに焦点を当てたがん治療
- 2019年11月号 チューブリンの過剰グルタミル化、ミトコンドリア、神経変性
- 2019年9月号 細胞運動性を制御するために相互作用する膜張力とアクチン細胞骨格
- 2019年8月号 Rac1B、がん、およびRac1
- 2019年7月号 Rhoファミリー GTPases、神経可塑性、およびうつ状態
- 2019年6月号 アクチンメチオニン酸化: 動的制御の次の段階
- 2019年5月号 ミクログリアと神経変性疾患
- 2019年2月号 生細胞画像化に対するCNS疾患や障害
2018年
- 2018年12月号 アクチン細胞骨格とメカノトランスダクション(機械的シグナル伝達)
- 2018年11月号 軸索再生と細胞骨格
- 2018年10月号 ニューロンにおける微小管と極性
- 2018年8月号 Rab GTPase と 神経変性
- 2018年7月号 SUMO レスリング: バランスが全て
- 2018年6月号 なぜ K-Ras は発がん特異性を示すのか?
- 2018年5月号 治療標的としてのユビキチンプロテアソームシステム:チューブリンは関与するか?
- 2018年4月号 RhoファミリーGEFと樹状突起スパインの構造的可塑性
- 2018年3月号 βカテニンとTFC/LEF-1の翻訳後修飾による標準的なWntシグナル制御
- 2018年2月号 がん抑制遺伝子p53の翻訳後修飾による機能の調整
- 2018年1月号 自閉スペクトラム症におけるGEF Trioの役割
2017年
- 2017年12月号 プロフィリン: アクチン結合タンパク質の多機能な役割
- 2017年11月号 ミトコンドリアにおけるアセチル化:新たな考え方と治療への応用の可能性
- 2017年9月号 翻訳後修飾のアセチル化による微小管の安定化
- 2017年8月号 神経軸索におけるアクチンリングを基盤とした周期的膜骨格(PMS)
- 2017年7月号 E3ユビキチンリガーゼMdm2によるがん抑制遺伝子p53の翻訳後制御
- 2017年6月号 多能性幹細胞(PSC)での転写因子による翻訳後制御
- 2017年5月号 Arf6 GEF と癌細胞の浸潤・転移
- 2017年4月号 PTEN(Phosphatase and Tensin Homolog)による翻訳後制御
- 2017年3月号 Tau の翻訳後修飾: アルツハイマー病の治療標的
- 2017年2月号 樹状細胞の移動におけるアクチン結合タンパク質とF-アクチン
2016年
- 2016年11月/12月号 GEF を介した GTPase シグナル伝達の低分子阻害剤
- 2016年9月号 FtsZ タンパク質: 抗菌薬の新規ターゲット
- 2016年7月号 翻訳後修飾(PTM)は心臓病において細胞骨格タンパク質を調節する
- 2016年6月号 モータータンパク質キネシンと神経変性
- 2016年5月号 チロシンリン酸化は Rhoファミリー GTPase 活性を調節する
- 2016年4月号 Rac1と糖尿病: ポジティブな役割とネガティブな役割
- 2016年3月号 SUMO化: 細胞骨格タンパク質の機能を調節するレギュレーター
- 2016年1月/2月号 ビメンチン中間径フィラメント: リン酸化による調節
2015年
- 2015年8月号 タンパク質調節に不可欠な翻訳後修飾
- 2015年7月号 アクチン細胞骨格のライブセルイメージング
- 2015年6月号 有糸分裂に関わるタンパク質のSUMO化: 局在と機能
- 2015年5月号 Ras 癌の治療: 5つの有望なターゲット
- 2015年4月号 Ras 依存性の癌で注目される YAP1
- 2015年3月号 増刊号 統合失調症において遺伝子変異により誘導されるアクチン依存のシナプスの変化
- 2015年3月号 Ral GTPase を調節する翻訳後修飾
- 2015年1月/2月号 RhoA のリン酸化はシグナル伝達を調節する
- 2015年1月号 増刊号 微小管を不安定化する suprafenacine: 新規抗癌剤のリード化合物としての可能性
2014年
- 2014年12月号 増刊号 RhoA は心筋細胞におけるアクチン細胞骨格の再構成とグルコース取り込みを仲介する
- 2014年11月号 増刊号 樹状突起の形態形成: ドーパミンD1受容体 および Rho ファミリー GTPase による制御
- 2014年11月/12月号 GTPase 活性化アッセイ: アイソフォームの検出
- 2014年10月号 アルギニンの正電荷を消失させるシトルリン化
- 2014年9月号 キネシンサブドメインの探索
- 2014年9月号 増刊号 アクチン結合タンパク質コフィリンの S-ニトロシル化: 細胞移動に対する影響
- 2014年8月号 増刊号 原発性硬化性胆管炎における N-Ras 発現および活性
- 2014年8月号 SUMO化: 細胞骨格タンパク質を標的とした翻訳後修飾
- 2014年7月号 Sos/K-Ras 結合を介して Ras シグナル伝達を制御する新しい低分子阻害剤
- 2014年6月号 増刊号 頭頸部扁平上皮癌における microRNA-138 による RhoC のダウンレギュレーション
- 2014年6月号 Rho GTPase と活性酸素種: クロストークとフィードバック
- 2014年5月号 ミオシンのアセチル化はサルコメアの構造と機能を調節する
- 2014年4月号 リジンのアセチル化 - 多様な細胞プロセスの制御因子
- 2014年3月号 インテグリンを介したβ-アクチンの酸化還元制御: PDIの出現
- 2014年1/2月号 ダイニン: 一つのモーターが関わる複数の神経変性疾患
2013年
- 2013年11/12月号 ダイニン:チームとして強力に作用するモータータンパク質
- 2013年10月号 神経変性:Rhes、SUMO化、ハンチントン病
- 2013年9月号 モノユビキチン化:タンパク質調節のダイナミックなタグ
- 2013年8月号 Ras及びRhoのプレニル化による翻訳後修飾:癌創薬における役割
- 2013年7月号 アクチンが引き起こす膜突起による浸潤:コルタクチン
- 2013年6月号 アクチン修飾と細胞骨格
- 2013年5月号 微小管内部の実体
- 2013年4月号 神経変性におけるTauの多面性
- 2013年3月号 蛍光フィブロネクチンタンパク質を用いた特発性肺線維症の創薬
- 2013年1/2月号 樹状突起棘:発生におけるArf6の役割
2012年
- 2012年11/12月号 ミオシンの小分子モジュレーター
- 2012年10月号 Rhoファミリーパスウェイのユビキチン化と制御
- 2012年9月号 神経変性におけるRac1 GTPaseの機能
- 2012年8月号 上皮間葉転換(EMT)とRhoファミリー低分子量G-タンパク質の関与
- 2012年7月号 チューブリンの多重修飾:グルタミル化とグリシル化
- 2012年6月号 細胞接着のフィブロネクチン制御と原線維形成
- 2012年5月号 アクチン酸化サイクルの機能
- 2012年4月号 トラフィッキング:ArfとCdc42/Racの結合
- 2012年3月号 G-LISAを用いた心臓研究: 糖尿病性心筋症におけるRho経路に関する研究
- 2012年1月/2月号 FtsZ: 新たな抗生剤の標的となるチュ−ブリンホモログ
商品は「研究用試薬」です。人や動物の医療用・臨床診断用・食品用としては使用しないように、十分ご注意ください。
※ 表示価格について














