






















 このページを印刷する
このページを印刷する
記事ID : 34705
ニューロンにおける微小管と極性 CYTOSKELETON NEWS 2018年10月号
ニューロンにおける微小管と極性
ニューロンの極性は、初期発生においてニューロンで生ずる空間的、形態的、構造的、および機能的分化を描写しており、これにより単一軸索と複数の樹状突起が形成されます。軸索と樹状突起は、シナプス後細胞の樹状突起からシナプス後ニューロンの軸索へと情報を受容し、処理し、伝播するといった、ニューロンにおける方向性をもったシグナル伝達において重要な役割を担います。樹状突起における興奮性インプットのほとんどは樹状突起棘で生じます。ニューロンの極性化は、少数の神経突起形成を経て円形の新生ニューロンがその対称性形状を損失するのと共に始まります1-4。ニューロンの極性化は、1) 細胞内の主要な細胞骨格ポリマーである微小管(MTs)の極性と、2) 軸索や樹状突起に存在するMTsに沿ってキネシンやダイニンによる極性化された積荷輸送に依存します4, 5。
MTsは、プラス端にβチューブリンが露出しマイナス端にαチューブリンが露出した、α/βチューブリンヘテロ二量体から構成される本質的に極性な繊維です5-7。MT極性により、1) MT会合/解体の位置、2) 細胞内のMT関連タンパク質(MAPs、例えば+TIPsモーター)がMTsに結合する場所、3) MTsに沿ってモーターに駆動される出入りが配向されます。重要なこととして、MTsは、ほぼ全ての正常神経細胞機能や複数の神経性病理が根底となるMT破壊に不可欠です7-10。
ニューロンにおける極性
軸索内では、MTsはプラス末端が均一に軸索末端(細胞体から遠位のプラス端)方向に向いた密接に束ねられたポリマーであるのに対し、樹状突起では、MTsは極性が混在しています(不均一に向いている)11(図1)。ダイニンモータータンパク質はMTsを軸索へと輸送し、これによりプラス端が遠位に配向されますが、キネシン-6はMTsを樹状突起へマイナス端遠位となるよう輸送します5。極性が混在したMTsは主に近位の樹状突起に存在し、プラス端が突出したMTsは樹状突起の遠位領域に存在します。樹状突起棘内では、遠位のプラス端が突出した方向性をもつ動的MTsは存在時間が短く、頭棘形態やシナプス可塑性および神経伝達に関与することが報告されています12, 13。伸長軸索の遠位部末端は成長円錐で、動的なプラス端遠位のチロシン化されたMTsから構成されています14。成長円錐内は区画化されており、MTsは中央(C) と末梢 (P) ドメインに存在します。前者は安定MTsを含み、後者は動的MTsを含んでいます。MTsに沿った順行性輸送が促進されることで、必要な分子やオルガネラが進行中の軸索成長円錐に提供されます4。成長円錐の発達時に末梢ドメイン中のMTs数が増加しますが、おそらく成長円錐を前進させるための力を産生する目的であると思われます4。末梢ドメインにおけるMTsの重合、脱重合、安定化、および不安定化は、アクチン細胞骨格との動的MTsの共役、Rac1介在性の腫瘍性タンパク質18/スタスミンの活性化、およびPI3K介在性のMAPsや+TIPs4群の活性化といった数々の制御力を受ける可能性があります。末梢ドメインのMTsは、軸索と樹状突起成長に重要な過程である膜内挿入に必須です。膜への挿入を介して、既存する膜の拡張表面領域上の張力が低減します。この張力減少がないと、膜突出、成長円錐の前進、およびニューロン極性化の継続は生じません。末梢ドメインのMTsの安定化により、これらMTsに沿ったシグナル伝達分子の輸送と必要な機械力が成長円錐中に産生します。安定化された末梢ドメインのMTsはアクチン動態、膜挿入時のアクチン介在性の力発生、および成長円錐の操縦と成長を調和させます4。
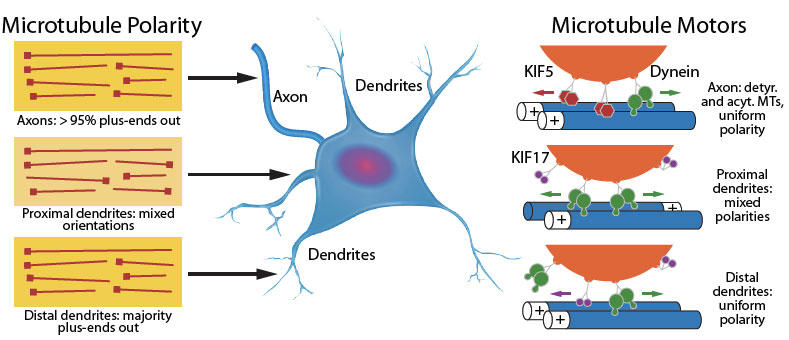
図.1 軸索と樹状突起におけるMT局在化。
MTの方向性は軸索においてほぼ全体的にプラス端突出であるのに対し、樹状突起では極性が混在しており、近位の樹状突起領域ではMTsのプラス端とマイナス端突出が混在し、遠位の樹状突起領域ではほとんどのMTsのプラス端が突出している。MTsに沿ったモーター介在性輸送は局在化している。軸索ではキネシンモーター(KIF5など)は積荷を順行性で輸送するのに対し、ダイニンは逆行性で輸送する。MTsの極性が混在している樹状突起の近位では、ダイニンは積荷を双方向性で輸送する。主にMTsのプラス端が突出している遠位の樹状突起では、KIF17が積荷を細胞体より離して輸送する。
PTMsとMTの極性
軸索のMTsと樹状突起のそれは翻訳後修飾(PTMs)により極性化されます。樹状突起では、混合極性をもつMTsがチロシン化、アセチル化、および短鎖グルタミン酸化により修飾を受けますが、ほとんどのマイナス端が突出したMTsは安定しアセチル化されるのに対し、プラス端突出MTsはチロシン化され動的です15。軸索のMTs(プラス端突出)は長鎖グルタミン酸化、アセチル化、ポリアミン化、脱チロシン化、およびΔ-2 チューブリンにより修飾されます14。
モーター極性
軸索と樹状突起内では、MTsは積荷がキネシンやダイニンモータータンパク質のより順行性または逆行性で輸送される際の軌道となります(図1)。少なくともキネシンの場合は、PTMsが特異的に修飾されたMTsへのキネシンの結合優先度に影響を及ぼします。例えば、キネシン-1はプラス端配向性モーターであり、アセチル化MTsと優先的に相互作用します。これにより、キネシン-1が樹状突起から出て軸索に入ります。逆に、キネシン-3はチロシン化MTsを好み、このキネシンモーターは軸索と樹状突起の双方で機能します15。キネシン介在性(例えばキネシン-1/KIF5とキネシン-2/KIF17など)輸送は、均一に方向付けられたプラス端突出MTsに沿って生ずるのに対し、ダイニンは極性が混在するMTsに沿った輸送を媒介します7, 16, 17。近位の樹状突起内の極性が混在したMTsは、双方向的なダイニン介在性積荷輸送を補助し、樹状突起の遠位部分に存在するプラス端突出MTsはKIF17-介在性輸送を利用します17(図1)。KIF5とKIF17は樹状突起に既に局在化している積荷を輸送することができ、さらに、タキソール安定化MTsは樹状突起へのKIF5介在性の積荷輸送を行うことができます17。
まとめ
ニューロンの極性はニューロンの適切な発生、成長、および生理機能に必須です。ニューロン内において、MTとモータータンパク質の極性はニューロンの極性を構築し維持する上で必要です。しかし、未だよくわかっていないこともあります5。
- なぜ細胞質分子によっては軸索内に存在するものの樹状突起には存在しないのか。
- MAPsはそれぞれの種類の神経突起において、どのようにして異なった区分化をされるのか。
- なぜニューロンには1つの軸索に対して複数の樹状突起があるのか。
Cytoskeleton, Inc.のサイエンティストは、このような問いを始め、その他の中枢神経系におけるMT機能に関する様々な疑問に対して研究している研究者の方々のお役にたてるような試薬をご用意しています。弊社では、アセチル化、チロシンリン酸化、ユビキチン化、およびSUMO化といった内在性PTMsのレベルを定量するSignal-Seeker Enrichment kitsを始めとした有用な試薬や、精製細胞骨格タンパク質(アクチン、チューブリン、低分子GTPアーゼ、キネシン、およびダイニンモーターなど)、また、これらタンパク質活性測定用の機能分析キットなどをご用意しています。
参考文献
- Caceres A. et al. 2012. Neuronal polarity: demarcation, growth and commitment. Curr. Opin. Cell Biol. 24, 547-553.
- Kapitein L.C. and Hoogenraad C.C. 2011. Which way to go. Cytoskeletal organization and polarized transport in neurons. Mol. Cell. Neurosci. 46, 9-20.
- Kapitein L.C. and Hoogenraad C.C. 2015. Building the neuronal microtubule cytoskeleton. Neuron. 87, 492-506.
- Schelski M. and Bradke F. 2017. Neuronal polarization: From spatiotemporal signaling to cytoskeletal dynamics. Mol. Cell. Neurosci. 84, 11-28.
- Baas P.W. and Lin S. 2011. Hooks and comets: The story of microtubule polarity orientation in the neuron. Dev. Neurobiol. 71, 403-418.
- Voelzmann A. et al. 2016. A conceptual view at microtubule plus end dynamics in neuronal axons. Brain Res. Bull. 126, 226-237.
- Chakraborti S. et al. 2016. The emerging role of the tubulin code: From the tubulin molecule to neuronal function and disease. Cytoskeleton. 73, 521-550.
- Tischfield M.A. et al. 2011. Phenotypic spectrum of the tubulin-related disorders and functional implications of disease-causing mutations. Curr. Opin. Genet. Dev. 21, 286-294.
- Adalbert R. and Coleman M.P. 2012. Axon pathology in age-related neurodegenerative disorders. Neuropathol. Appl. Neurobiol. 39, 90-108.
- Neumann B. and Hilliar M.A. 2014. Loss of MEC-17 leads to microtubule instability and axonal degeneration. Cell Rep. 6, 93-103.
- Coles C.H. and Bradke F. 2015. Coordinating neuronal actin-microtubule dynamics. Curr. Biol. 25, R677-R691.
- Hu X. et al. 2008. Activity-dependent dynamic microtubule invasion of dendritic spines. J. Neurosci. 28, 13094-13105.
- Jaworski J. et al. 2009. Dynamic microtubules regulate dendritic spine morphology and synaptic plasticity. Neuron. 61, 85-100.
- Park J.H. and Roll-Mecak A. 2018. The tubulin code in neuronal polarity. Curr. Opin. Neurobiol. 51, 95-102.
- Tas R.P. et al. 2017. Differentiation between oppositely oriented microtubules controls polarized neuronal transport. Neuron. 96, 1264-1271.
- Prokop A. 2013. The intricate relationship between microtubules and their associated motor proteins during axon growth and maintenance. Neural Dev. 8, 17.
- Kapitein L.C. et al. 2010. Mixed microtubules steer dynein-driven cargo transport into dendrites. Curr. Biol. 20, 290-299.
Signal Seeker™ Kits
[商品詳細]
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 | 在庫 |
|---|---|---|---|---|---|
Signal-SeekerTM Acetyl-Lysine Detection Kit |
CYT | BK163 | 30 ASSAY |
CYT社 BK163L 1 を参照 |
なし |
| Signal-SeekerTM Phosphotyrosine Detection Kit |
CYT | BK160 | 30 ASSAY |
CYT社 BK160L 1 を参照 |
なし |
| Signal-SeekerTM Ubiquitin Detection Kit |
CYT | BK161 | 30 ASSAY |
CYT社 BK161L 1 を参照 |
なし |
| Signal-SeekerTM SUMOylation Detection Kit |
CYT | BK162 | 30 ASSAY |
CYT社 BK162L 1 を参照 |
なし |
Tubulin and Kinesin Reagents
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 | 在庫 |
|---|---|---|---|---|---|
| SiR-Tubulin Kit |
CYT | CY-SC002 | 1 KIT [50-300 slides] |
¥182,000 | なし |
| Cytoplasmic Dynein Motor Protein, Porcine |
CYT | CS-DN01 | 1*50 UG |
¥153,000 | なし |
Tubulin Kits
[商品詳細]
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 | 在庫 |
|---|---|---|---|---|---|
| Tubulin polymerization assay using >99% pure tubulin - OD based, Porcine |
CYT | BK006P | 1 KIT [24 assays] |
¥300,000 | なし |
| Tubulin polymerization assay using >99% pure tubulin - fluorescence based |
CYT | BK011P | 1 KIT [96 assays] |
¥317,000 | なし |
| Microtubule Binding Protein Spin-Down Assay Biochem Kit |
CYT | BK029 | 1 KIT [30-100 assays] |
¥265,000 | なし |
ATPase/GTPase Kits
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 | 在庫 |
|---|---|---|---|---|---|
| ATPase ELIPATM (Enzyme Linked Inorganic Phosphate Assay) |
CYT | BK051 | 1 KIT [96 assays] |
CYT社 BK05152 1 を参照 |
なし |
| GTPase ELIPATM (Enzyme Linked Inorganic Phosphate Assay) |
CYT | BK052 | 1 KIT [96 assays] |
CYT社 BK05152 1 を参照 |
なし |
| HTS kinesin ATPase Endpoint Assay Biochem Kit |
CYT | BK053 | 1 KIT [1000 assays] |
¥307,000 | なし |
| CytoPhos Endpoint Phosphate Assay |
CYT | BK054 | 1 KIT [1000 assays] |
¥134,000 | なし |
| Kinesin ELIPA kit |
CYT | BK060 | 1 KIT [96 assays] |
¥278,000 | なし |
■ CYTOSKELETON NEWS バックナンバー
- 2020年10月号 紡錘体 - 可視化に向けた新規ツール
- 2020年8月号 細胞膜染色用蛍光プローブ
- 2020年4月号 生細胞におけるF-アクチンプローブ
- 2020年3月号 コロナウイルスと細胞骨格
- 2020年2月号 タウ(Tau)の将来性をMapping
- 2020年1月号 Rho GTPaseによる細胞遊走制御
- 2019年12月号 表現型プロファイリング:アクチンに焦点を当てたがん治療
- 2019年11月号 チューブリンの過剰グルタミル化、ミトコンドリア、神経変性
- 2019年9月号 細胞運動性を制御するために相互作用する膜張力とアクチン細胞骨格
- 2019年8月号 Rac1B、がん、およびRac1
- 2019年7月号 Rhoファミリー GTPases、神経可塑性、およびうつ状態
- 2019年6月号 アクチンメチオニン酸化: 動的制御の次の段階
- 2019年5月号 ミクログリアと神経変性疾患
- 2019年2月号 生細胞画像化に対するCNS疾患や障害
2018年
- 2018年12月号 アクチン細胞骨格とメカノトランスダクション(機械的シグナル伝達)
- 2018年11月号 軸索再生と細胞骨格
- 2018年10月号 ニューロンにおける微小管と極性
- 2018年8月号 Rab GTPase と 神経変性
- 2018年7月号 SUMO レスリング: バランスが全て
- 2018年6月号 なぜ K-Ras は発がん特異性を示すのか?
- 2018年5月号 治療標的としてのユビキチンプロテアソームシステム:チューブリンは関与するか?
- 2018年4月号 RhoファミリーGEFと樹状突起スパインの構造的可塑性
- 2018年3月号 βカテニンとTFC/LEF-1の翻訳後修飾による標準的なWntシグナル制御
- 2018年2月号 がん抑制遺伝子p53の翻訳後修飾による機能の調整
- 2018年1月号 自閉スペクトラム症におけるGEF Trioの役割
2017年
- 2017年12月号 プロフィリン: アクチン結合タンパク質の多機能な役割
- 2017年11月号 ミトコンドリアにおけるアセチル化:新たな考え方と治療への応用の可能性
- 2017年9月号 翻訳後修飾のアセチル化による微小管の安定化
- 2017年8月号 神経軸索におけるアクチンリングを基盤とした周期的膜骨格(PMS)
- 2017年7月号 E3ユビキチンリガーゼMdm2によるがん抑制遺伝子p53の翻訳後制御
- 2017年6月号 多能性幹細胞(PSC)での転写因子による翻訳後制御
- 2017年5月号 Arf6 GEF と癌細胞の浸潤・転移
- 2017年4月号 PTEN(Phosphatase and Tensin Homolog)による翻訳後制御
- 2017年3月号 Tau の翻訳後修飾: アルツハイマー病の治療標的
- 2017年2月号 樹状細胞の移動におけるアクチン結合タンパク質とF-アクチン
2016年
- 2016年11月/12月号 GEF を介した GTPase シグナル伝達の低分子阻害剤
- 2016年9月号 FtsZ タンパク質: 抗菌薬の新規ターゲット
- 2016年7月号 翻訳後修飾(PTM)は心臓病において細胞骨格タンパク質を調節する
- 2016年6月号 モータータンパク質キネシンと神経変性
- 2016年5月号 チロシンリン酸化は Rhoファミリー GTPase 活性を調節する
- 2016年4月号 Rac1と糖尿病: ポジティブな役割とネガティブな役割
- 2016年3月号 SUMO化: 細胞骨格タンパク質の機能を調節するレギュレーター
- 2016年1月/2月号 ビメンチン中間径フィラメント: リン酸化による調節
2015年
- 2015年8月号 タンパク質調節に不可欠な翻訳後修飾
- 2015年7月号 アクチン細胞骨格のライブセルイメージング
- 2015年6月号 有糸分裂に関わるタンパク質のSUMO化: 局在と機能
- 2015年5月号 Ras 癌の治療: 5つの有望なターゲット
- 2015年4月号 Ras 依存性の癌で注目される YAP1
- 2015年3月号 増刊号 統合失調症において遺伝子変異により誘導されるアクチン依存のシナプスの変化
- 2015年3月号 Ral GTPase を調節する翻訳後修飾
- 2015年1月/2月号 RhoA のリン酸化はシグナル伝達を調節する
- 2015年1月号 増刊号 微小管を不安定化する suprafenacine: 新規抗癌剤のリード化合物としての可能性
2014年
- 2014年12月号 増刊号 RhoA は心筋細胞におけるアクチン細胞骨格の再構成とグルコース取り込みを仲介する
- 2014年11月号 増刊号 樹状突起の形態形成: ドーパミンD1受容体 および Rho ファミリー GTPase による制御
- 2014年11月/12月号 GTPase 活性化アッセイ: アイソフォームの検出
- 2014年10月号 アルギニンの正電荷を消失させるシトルリン化
- 2014年9月号 キネシンサブドメインの探索
- 2014年9月号 増刊号 アクチン結合タンパク質コフィリンの S-ニトロシル化: 細胞移動に対する影響
- 2014年8月号 増刊号 原発性硬化性胆管炎における N-Ras 発現および活性
- 2014年8月号 SUMO化: 細胞骨格タンパク質を標的とした翻訳後修飾
- 2014年7月号 Sos/K-Ras 結合を介して Ras シグナル伝達を制御する新しい低分子阻害剤
- 2014年6月号 増刊号 頭頸部扁平上皮癌における microRNA-138 による RhoC のダウンレギュレーション
- 2014年6月号 Rho GTPase と活性酸素種: クロストークとフィードバック
- 2014年5月号 ミオシンのアセチル化はサルコメアの構造と機能を調節する
- 2014年4月号 リジンのアセチル化 - 多様な細胞プロセスの制御因子
- 2014年3月号 インテグリンを介したβ-アクチンの酸化還元制御: PDIの出現
- 2014年1/2月号 ダイニン: 一つのモーターが関わる複数の神経変性疾患
2013年
- 2013年11/12月号 ダイニン:チームとして強力に作用するモータータンパク質
- 2013年10月号 神経変性:Rhes、SUMO化、ハンチントン病
- 2013年9月号 モノユビキチン化:タンパク質調節のダイナミックなタグ
- 2013年8月号 Ras及びRhoのプレニル化による翻訳後修飾:癌創薬における役割
- 2013年7月号 アクチンが引き起こす膜突起による浸潤:コルタクチン
- 2013年6月号 アクチン修飾と細胞骨格
- 2013年5月号 微小管内部の実体
- 2013年4月号 神経変性におけるTauの多面性
- 2013年3月号 蛍光フィブロネクチンタンパク質を用いた特発性肺線維症の創薬
- 2013年1/2月号 樹状突起棘:発生におけるArf6の役割
2012年
- 2012年11/12月号 ミオシンの小分子モジュレーター
- 2012年10月号 Rhoファミリーパスウェイのユビキチン化と制御
- 2012年9月号 神経変性におけるRac1 GTPaseの機能
- 2012年8月号 上皮間葉転換(EMT)とRhoファミリー低分子量G-タンパク質の関与
- 2012年7月号 チューブリンの多重修飾:グルタミル化とグリシル化
- 2012年6月号 細胞接着のフィブロネクチン制御と原線維形成
- 2012年5月号 アクチン酸化サイクルの機能
- 2012年4月号 トラフィッキング:ArfとCdc42/Racの結合
- 2012年3月号 G-LISAを用いた心臓研究: 糖尿病性心筋症におけるRho経路に関する研究
- 2012年1月/2月号 FtsZ: 新たな抗生剤の標的となるチュ−ブリンホモログ
商品は「研究用試薬」です。人や動物の医療用・臨床診断用・食品用としては使用しないように、十分ご注意ください。
※ 表示価格について














