






















 このページを印刷する
このページを印刷する
記事ID : 35949
アクチンメチオニン酸化: 動的制御の次の段階 CYTOSKELETON NEWS 2019年6月号
アクチンPTMの背景
アクチンは、十分に特徴づけられた、豊富に発現している必須の細胞骨格タンパク質です。その動的特性により単量体(G-アクチン)と重合体(F-アクチン)状態とを移行することができ、これは数々の細胞プロセスにおいて非常に重要です。アクチンの動態や機能は数々の内部刺激と外部刺激により制御されており、これらはアクチン結合タンパク質(ABPs)、シグナル伝達物質、および他のタンパク質により促進されます。また、現在ではいくつかの研究より、アクチン自体が翻訳後修飾(PTMs)により高度に修飾されていることが示唆されています。その上、特定アクチンのPTMsに特化した研究により、そのアクチン動態、ABP相互作用、およびアクチン依存性生理機能への影響について詳述されました 1,2)。例えば、アクチンN-末端アセチル化、リジンアセチル化、アルギニル化、SUMO化、およびユビキチン化を研究されてきました。ここでは、メチオニン(Met)44とMet47におけるアクチンの生理的酸化に関する現在の研究を紹介いたします。
アクチン酸化:メチオニンスルホキシドに焦点を当てる(MetO)
酸化分野では一般的にH2O2といった自然に形成された酸化体により誘導される病理学的酸化(酸化ストレス)に焦点をあてており、最初は、酸化は“有毒”であると仮説を立てられていたものの、現在では酸化は病理学的現象と生理学的現象の双方にとってのシグナル伝達機構であることが知られています 3)。活性酸素種(ROS)に関する研究のほとんどはチオールを基盤としたシステイン(Cys)修飾に焦点を当てています。これはアクチンに関しても当てはまるため、アクチン酸化に関する初期の研究の大部分ではH2O2処理を利用していました。これより、亢進したラグタイム、重合速度の遅延、重合レベルの低下によって明示されるようにF-アクチン含量や重合活性の変化といった結果が得られていました 4-6)。さらなる研究より、H2O2誘導性アクチン酸化はin vitroにおいて最初にCys374を標的とするものの、Met44、Met47、Met176、Met190、Met269、およびMet355を始めとするいくつかのMetを標的することが示されています 7)。MetO酸化は、in vivoにおいて正常条件またはストレス条件下において生じます。しかし、Manta and Gladsyshevらは、ROS誘導性MetO形成はin vivoにおいてMsrAなどの還元酵素の速度式に比べ非効率的に生ずることを報告しています 8)。したがって、in vivoにおいてアクチンのメチオニンが酵素的に酸化するのかどうかといった疑問は未だに解決されていません。Termanらによる精液の研究より、in vitroにおいて、酵素であるMICAL(CasL関連分子)のアクチンのMet44とMet47の酸化における役割が同定されました 9)。このin vitro研究より、線維切断や重合低減の両方を促進するためにはMet44におけるMetOで十分であることが示されました。さらに、ショウジョウバエにおいてMICALの過剰発現によりbristle formationが変形するもののM44Lアクチン変異体では低減するように、アクチンMetOの誤制御により重度の形態学上の影響が生ずることから、in vivoにおけるその役割が明確となりました 9,10)。近年の研究より、この機構が細胞質分裂の終末段階におけるアクチン脱重合化に必須であることが特定され、F-アクチンのMICAL-1酸化の生理学的役割が同定されました 11)。これらの研究により、アクチンの分子的機能、細胞機能、形態的機能に対するアクチンMetOのもつ生理学的重要性に光が投じられました。
アクチン酸化の制御機構
MICALは細胞内フラボタンパク質モノオキシゲナーゼであり、昆虫から哺乳動物にかけて保存されており、酸化還元(redox)反応の触媒として機能します 12)。哺乳動物では3種のMICALファミリメンバーが存在し、この全てがアクチンを酸化できるもののそれぞれ異なった動態を示し、空間的局在化や制御を行います 13)。Termanのグループは、MICALがアクチンと相互作用し、NADPHを補助因子としてアクチンをMet44とMet479において酸化することを示しました。Met44残基はアクチンのサブドメイン2のD-ループに存在し、これはアクチンサブユニット接触に必須です 14)。Met44は酸化されると、負に帯電してアクチン単量体間相互作用と干渉します。そのため、F-アクチン切断と脱重合が促進します。重要なこととして、DTTのような還元体はMICAL活性を変えないため、アクチンへのMICAL介在性効果はH2O2といった拡散性酸化体を通しては生じず、MICALはアクチンに非常に接近している必要があります 9)。
近年の研究より、MICAL介在性のアクチンMetOはメチオニンスルホキシド還元酵素のSelR/MsrBファミリーにより反転することが発見されました。2つの研究グループがそれぞれ独自に、SelR (MsrB) がMet44とMet47の還元や正常アクチン動態の回復に原因となる酵素であることを同定しました 15,16)。これらの研究より、SelR/MsrBが選択的にMICALにより酸化されたアクチンを還元し、他のメチオニンスルホキシド還元酵素メンバーであるMsrAは関与しないことがわかりました。SelR/MsrBは特異的にR-異性体MetOを還元し、MsrAは特異的にS-異性体MetOを還元することから、彼らはMetO R-異性体と共にMICAL立体特異的にアクチンを酸化すると結論付けています。以上より、これらのデータはアクチンの特異的MetO制御を介したアクチン動態と細胞骨格形成を制御する、可逆的で特異的な酸化還元システムを説明するものと考えられます。
アクチン酸化の生理学
MICALsは遍在的に発現し、アクチンのMet44とMet47残基は非常に保存されていることから、この酸化還元制御の機構が全ての組織や細胞型においてアクチン機能を調節する上で卓越した役割を担うものと思われます。いくつかの研究では、MICALタンパク質が一連の組織や生物において生理学的にも病理学的にも役割を担うことに着目しています。しかし、これらの効果がアクチン依存性制御を介して促進しているか否かは完全には研究されていません 17)。以下にMICALがその特異的なアクチン制御に関連して有する著しい効果の例をいくつ挙げました。例えば、このような研究の一つに、MICAL-2を核のG-アクチンレベル制御因子として同定し、続いてMRTF-A/SRF転写制御、およびゼブラフィッシュにおける心臓発生の生理学的制御として同定したものがあります 18)。その他に、増殖因子や化学忌避物質がアクチンのMICAL制御に及ぼす影響を調査したものがあり、その結果、逆説的に化学忌避物質の効果が増殖因子シグナル伝達により増幅され、これが病理学的腫瘍増殖や治療への応答と同様に生理学的軸索ガイダンス制御へ顕著な影響を及ぼすことを見出しました 19)。最後に、MICAL-1はF-アクチン制御を介した海馬苔状線維結合の発生に重要であり、この生理学的プロセスは神経以上において重要となる可能性があります 20)。これらの研究などから、いくつかの特徴的な細胞プロセスにこのアクチンの小さな生理的MetOがもつ計り知れない効果に光が当てられています。
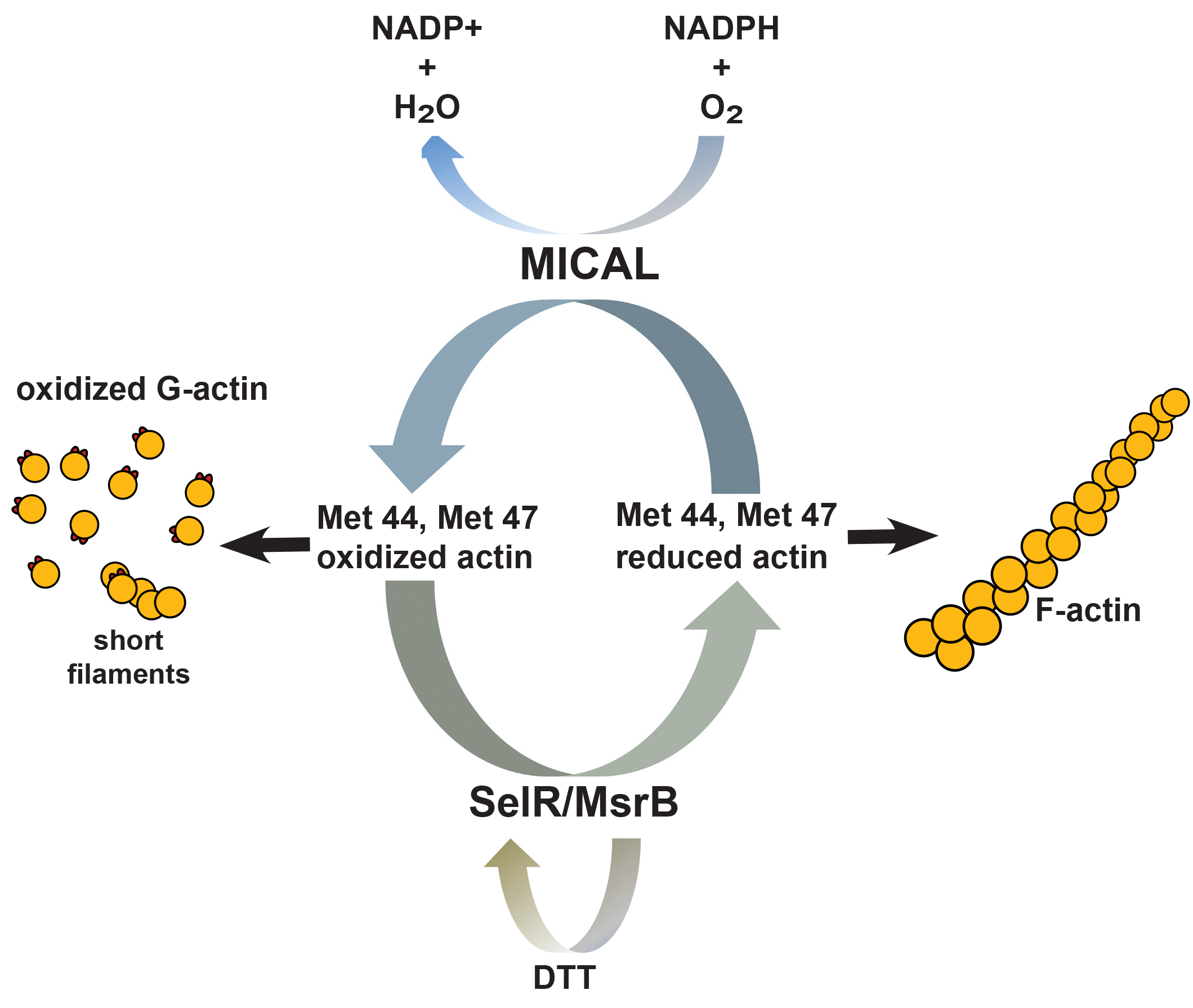
図1 アクチンMet44とMet47生理学的酸化還元システム
まとめ
これらの研究より、この新規の可逆的酸化還元制御機構の同定を介して確実にアクチンPTM分野の発展に向けて著しい基礎を築くことでしょう。近年の研究より、アクチンのMICAL制御は多くのアクチン依存性細胞プロセスにおいて重要であると思われ、アクチン生物学においてこの酸化還元機構がどれだけ公汎しているか驚くことでしょう。同様に、アクチンのABP制御がいかに共同して機能するのか、また、アクチンMetOのように決定的なPTMsとは反対に機能するのかを特定することは興味深いことでしょう。近年の研究より、アクチン酸化とコフィリンが共同してアクチンを分解すること 21)が示唆されており、これはABPとアクチンPTMとの掛け合い応答が存在することを示しており、更なる検討を行う正当性が示されたといえます。研究者が疾患におけるMICAL酸化アクチンの役割を解読するにつれ、ROSと酵素的アクチンMetOとの相互作用をさらに検討することは興味深いことといえます。これらの種の疑問に取り組むために有用なMetOアクチンツールを有することで、研究者がアクチン生理学やMetOアクチンが各々の研究モデルにおいて役割を担うかどうかをよりよく理解することができるでしょう。Cytoskeleton社では、様々なMICAL-酸化型(MOX)アクチンツールをご用意しており、アクチン生物学に対してよりよく理解できるよう、この新規アクチン制御機構をご自身の研究へと組込むことをお手伝いしています。
参考文献
- Varland S. et al. 2019. Actin post-translational modifications: The Cinderella of cytoskeletal control. Trends Biochem. Sci. DOI: 10.1016/j.tibs.2018.11.010.
- Terman J.R. and Kashina A. 2013. Post-translational modification and regulation of actin. Curr. Opin. Cell Biol. 25, 30-38.
- Wilson C. and Gonzalez-Billault C. 2015. Regulation of cytoskeletal dynamics by redox signaling and oxidative stress: implications for neuronal development and trafficking. Front. Cell Neurosci. 9, 381.
- Hinshaw D.B. et al. 1986. Cytoskeletal and morphologic impact of cellular oxidant injury. Am. J. Pathol. 123, 454-464.
- DalleDonne I. et al. 1995. H2O2-treated actin: assembly and polymer interactions with cross-linking proteins. Biophys. J. 69, 2710-2719.
- Lassing I. et al. 2007. Molecular and structural basis for redox regulation of beta-actin. J. Mol. Biol. 370, 331-348.
- Milzani A. et al. 2000. The oxidation produced by hydrogen peroxide on Ca-ATP-G-actin. Protein Sci. 9, 1774-1782.
- Manta B. and Gladyshev V.N. 2017. Regulated methionine oxidation by monooxygenases. Free Rad. Biol. Med. 109, 141-155.
- Hung R.J. et al. 2011. Direct redox regulation of F-actin assembly and disassembly by Mical. Science. 334, 1710-1713.
- Hung R.J. et al. 2010. Mical links semaphorins to F-actin disassembly. Nature. 463, 823-827.
- Fremont S. et al. 2017. Oxidation of F-actin controls the terminal steps of cytokinesis. Nat. Commun. 8, 14528.
- Terman J.R. et al. 2002. MICALs, a family of conserved flavoprotein oxidoreductases, function in plexin-mediated axonal repulsion. Cell. 109, 887-900.
- Wu H. et al. 2018. The MICALs are a family of F-actin dismantling oxidoreductases conserved from Drosophila to humans. Sci. Rep. 8, 937.
- Grintsevich E.E. et al. 2017. Catastrophic disassembly of actin filaments via Mical-mediated oxidation. Nat. Commun. 8, 2183.
- Lee B.C. et al. 2013. MsrB1 and MICALs regulate actin assembly and macrophage function via reversible stereoselective methionine oxidation. Mol Cell. 51, 397-404.
- Hung R.J. et al. 2013. SelR reverses Mical-mediated oxidation of actin to regulate F-actin dynamics. Nat. Cell Biol. 15, 1445-1454.
- Wilson C. et al. 2016. Actin filaments-A target for redox regulation. Cytoskeleton (Hoboken). 73, 577-595.
- Lundquist M.R. et al. 2014. Redox modification of nuclear actin by MICAL-2 regulates SRF signaling. Cell. 156, 563-576.
- Yoon J. et al. 2017. Amplification of F-Actin disassembly and cellular repulsion by growth factor signaling. Dev. Cell. 42, 117-129.
- Van Battum E.Y. et al. 2014. The intracellular redox protein MICAL-1 regulates the development of hippocampal mossy fibre connections. Nat. Commun. 5, 4317.
- Grintsevich E.E. et al. 2016. F-actin dismantling through a redox-driven synergy between Mical and cofilin. Nat. Cell Biol. 18, 876-885.
MOX Actin Products
[商品詳細]
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 | 在庫 |
|---|---|---|---|---|---|
Actin, Rabbit, Unlabeled |
CYT | AKL99-A | 4*250 UG |
¥73,000 | なし |
| Actin, Rabbit, Unlabeled |
CYT | AKL99-B | 2*1 MG |
¥87,000 | なし |
| Actin, Rabbit, Unlabeled |
CYT | AKL99-C | 5*1 MG |
¥177,000 | なし |
| Actin, Rabbit, Unlabeled |
CYT | AKL99-D | 10*1 MG |
¥335,000 | なし |
| Actin, Rabbit, Unlabeled |
CYT | AKL99-E | 20*1 MG |
お問い合わせ | なし |
| MICAL-Oxidized (Pyrene labeled) Actin Protein, Rabbit |
CYT | MXAP95 | 250 UG |
販売終了 | 販売終了 |
| MICAL-Oxidized Actin Protein, Rabbit |
CYT | MXA95 | 250 UG |
CYT社 MXA95 2*250 を参照 |
なし |
| MICAL-1 Protein 6xHis, Human |
CYT | MIC01 | 2*50 UG |
CYT社 MIC01A 2*50 を参照 |
なし |
| MsrB2 Protein 6xHis, Human |
CYT | MB201 | 2*50 UG |
CYT社 MB201A 2*50 を参照 |
なし |
Actin Products
[商品詳細]
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 | 在庫 |
|---|---|---|---|---|---|
| Actin, Bovine, Unlabeled |
CYT | AD99-A | 1*1 MG |
¥44,000 | なし |
| Actin, Bovine, Unlabeled |
CYT | AD99-B | 5*1 MG |
¥181,000 | なし |
| Actin, Chicken |
CYT | AS99-A | 1*1 MG |
¥44,000 | なし |
| Actin, Chicken |
CYT | AS99-B | 5*1 MG |
¥181,000 | なし |
| Actin protein (pre-formed filaments), Rabbit |
CYT | AKF99-A | 1*1 MG |
¥69,000 | なし |
| Actin protein (pre-formed filaments), Rabbit |
CYT | AKF99-B | 5*1 MG |
¥240,000 | なし |
| Actin, Rabbit, Unlabeled |
CYT | AKL95-B | 1*1 MG |
¥35,000 | なし |
| Actin, Rabbit, Unlabeled |
CYT | AKL95-C | 5*1 MG |
¥137,000 | なし |
| Actin, Rabbit, Unlabeled |
CYT | AKL99-A | 4*250 UG |
¥73,000 | なし |
| Actin, Rabbit, Unlabeled |
CYT | AKL99-B | 2*1 MG |
¥87,000 | なし |
| Actin, Rabbit, Unlabeled |
CYT | AKL99-C | 5*1 MG |
¥177,000 | なし |
| Actin, Rabbit, Unlabeled |
CYT | AKL99-D | 10*1 MG |
¥335,000 | なし |
| Actin, Rabbit, Unlabeled |
CYT | AKL99-E | 20*1 MG |
お問い合わせ | なし |
| Actin, Human, Unlabeled |
CYT | APHL99-A | 2*250 UG |
¥79,000 | なし |
| Actin, Human, Unlabeled |
CYT | APHL99-C | 1*1 MG |
¥133,000 | なし |
| Actin, Human, Unlabeled |
CYT | APHL99-E | 5*1 MG |
お問い合わせ | なし |
| Actin, Human, Rhodamine Isothiocyanate |
CYT | APHR-A | 4*10 UG |
¥108,000 | なし |
| Actin, Human, Rhodamine Isothiocyanate |
CYT | APHR-C | 20*10 UG |
¥339,000 | なし |
| Actin, Rabbit, Rhodamine Isothiocyanate |
CYT | AR05-B | 10*20 UG |
¥104,000 | なし |
| Actin, Rabbit, Rhodamine Isothiocyanate |
CYT | AR05-C | 20*20 UG |
¥200,000 | なし |
| SiR-Actin Kit |
CYT | CY-SC001 | 1 KIT [50-300 slides] |
¥182,000 | なし |
| SiR700-Actin Kit |
CYT | CY-SC013 | 1 KIT [35-200 slides] |
¥182,000 | なし |
| Acti-stain 488 phalloidin, Plant |
CYT | PHDG1-A | 1*500 UL [300 slides] |
¥79,000 | あり |
| Acti-stain 555 phalloidin, Mushroom |
CYT | PHDH1-A | 1*500 UL [300 slides] |
¥79,000 | あり |
| Acti-stain 670 phalloidin, Plant |
CYT | PHDN1-A | 1*500 UL [300 slides] |
¥79,000 | あり |
| Rhodamine Phalloidin, Rhodamine Isothiocyanate |
CYT | PHDR1 | 1*500 UL [300 slides] |
¥79,000 | あり |
Actin Biochem Kits
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 | 在庫 |
|---|---|---|---|---|---|
| Actin Binding Protein Spin-Down Assay Biochem Kit, Rabbit |
CYT | BK001 | 1 KIT [30-100 assays] |
¥268,000 | なし |
| Actin Binding Protein Spin-Down Assay Biochem Kit, Human |
CYT | BK013 | 1 KIT [30-100 assays] |
¥272,000 | なし |
| Actin Polymerization Biochem Kit (fluorescence format), Rabbit |
CYT | BK003 | 1 KIT [30-100 assays] |
¥304,000 | なし |
| G-Actin/F-actin In Vivo Assay Biochem Kit, Mouse |
CYT | BK037 | 1 KIT [30-100 assays] |
¥264,000 | なし |

■ CYTOSKELETON NEWS バックナンバー
- 2020年10月号 紡錘体 - 可視化に向けた新規ツール
- 2020年8月号 細胞膜染色用蛍光プローブ
- 2020年4月号 生細胞におけるF-アクチンプローブ
- 2020年3月号 コロナウイルスと細胞骨格
- 2020年2月号 タウ(Tau)の将来性をMapping
- 2020年1月号 Rho GTPaseによる細胞遊走制御
- 2019年12月号 表現型プロファイリング:アクチンに焦点を当てたがん治療
- 2019年11月号 チューブリンの過剰グルタミル化、ミトコンドリア、神経変性
- 2019年9月号 細胞運動性を制御するために相互作用する膜張力とアクチン細胞骨格
- 2019年8月号 Rac1B、がん、およびRac1
- 2019年7月号 Rhoファミリー GTPases、神経可塑性、およびうつ状態
- 2019年6月号 アクチンメチオニン酸化: 動的制御の次の段階
- 2019年5月号 ミクログリアと神経変性疾患
- 2019年2月号 生細胞画像化に対するCNS疾患や障害
2018年
- 2018年12月号 アクチン細胞骨格とメカノトランスダクション(機械的シグナル伝達)
- 2018年11月号 軸索再生と細胞骨格
- 2018年10月号 ニューロンにおける微小管と極性
- 2018年8月号 Rab GTPase と 神経変性
- 2018年7月号 SUMO レスリング: バランスが全て
- 2018年6月号 なぜ K-Ras は発がん特異性を示すのか?
- 2018年5月号 治療標的としてのユビキチンプロテアソームシステム:チューブリンは関与するか?
- 2018年4月号 RhoファミリーGEFと樹状突起スパインの構造的可塑性
- 2018年3月号 βカテニンとTFC/LEF-1の翻訳後修飾による標準的なWntシグナル制御
- 2018年2月号 がん抑制遺伝子p53の翻訳後修飾による機能の調整
- 2018年1月号 自閉スペクトラム症におけるGEF Trioの役割
2017年
- 2017年12月号 プロフィリン: アクチン結合タンパク質の多機能な役割
- 2017年11月号 ミトコンドリアにおけるアセチル化:新たな考え方と治療への応用の可能性
- 2017年9月号 翻訳後修飾のアセチル化による微小管の安定化
- 2017年8月号 神経軸索におけるアクチンリングを基盤とした周期的膜骨格(PMS)
- 2017年7月号 E3ユビキチンリガーゼMdm2によるがん抑制遺伝子p53の翻訳後制御
- 2017年6月号 多能性幹細胞(PSC)での転写因子による翻訳後制御
- 2017年5月号 Arf6 GEF と癌細胞の浸潤・転移
- 2017年4月号 PTEN(Phosphatase and Tensin Homolog)による翻訳後制御
- 2017年3月号 Tau の翻訳後修飾: アルツハイマー病の治療標的
- 2017年2月号 樹状細胞の移動におけるアクチン結合タンパク質とF-アクチン
2016年
- 2016年11月/12月号 GEF を介した GTPase シグナル伝達の低分子阻害剤
- 2016年9月号 FtsZ タンパク質: 抗菌薬の新規ターゲット
- 2016年7月号 翻訳後修飾(PTM)は心臓病において細胞骨格タンパク質を調節する
- 2016年6月号 モータータンパク質キネシンと神経変性
- 2016年5月号 チロシンリン酸化は Rhoファミリー GTPase 活性を調節する
- 2016年4月号 Rac1と糖尿病: ポジティブな役割とネガティブな役割
- 2016年3月号 SUMO化: 細胞骨格タンパク質の機能を調節するレギュレーター
- 2016年1月/2月号 ビメンチン中間径フィラメント: リン酸化による調節
2015年
- 2015年8月号 タンパク質調節に不可欠な翻訳後修飾
- 2015年7月号 アクチン細胞骨格のライブセルイメージング
- 2015年6月号 有糸分裂に関わるタンパク質のSUMO化: 局在と機能
- 2015年5月号 Ras 癌の治療: 5つの有望なターゲット
- 2015年4月号 Ras 依存性の癌で注目される YAP1
- 2015年3月号 増刊号 統合失調症において遺伝子変異により誘導されるアクチン依存のシナプスの変化
- 2015年3月号 Ral GTPase を調節する翻訳後修飾
- 2015年1月/2月号 RhoA のリン酸化はシグナル伝達を調節する
- 2015年1月号 増刊号 微小管を不安定化する suprafenacine: 新規抗癌剤のリード化合物としての可能性
2014年
- 2014年12月号 増刊号 RhoA は心筋細胞におけるアクチン細胞骨格の再構成とグルコース取り込みを仲介する
- 2014年11月号 増刊号 樹状突起の形態形成: ドーパミンD1受容体 および Rho ファミリー GTPase による制御
- 2014年11月/12月号 GTPase 活性化アッセイ: アイソフォームの検出
- 2014年10月号 アルギニンの正電荷を消失させるシトルリン化
- 2014年9月号 キネシンサブドメインの探索
- 2014年9月号 増刊号 アクチン結合タンパク質コフィリンの S-ニトロシル化: 細胞移動に対する影響
- 2014年8月号 増刊号 原発性硬化性胆管炎における N-Ras 発現および活性
- 2014年8月号 SUMO化: 細胞骨格タンパク質を標的とした翻訳後修飾
- 2014年7月号 Sos/K-Ras 結合を介して Ras シグナル伝達を制御する新しい低分子阻害剤
- 2014年6月号 増刊号 頭頸部扁平上皮癌における microRNA-138 による RhoC のダウンレギュレーション
- 2014年6月号 Rho GTPase と活性酸素種: クロストークとフィードバック
- 2014年5月号 ミオシンのアセチル化はサルコメアの構造と機能を調節する
- 2014年4月号 リジンのアセチル化 - 多様な細胞プロセスの制御因子
- 2014年3月号 インテグリンを介したβ-アクチンの酸化還元制御: PDIの出現
- 2014年1/2月号 ダイニン: 一つのモーターが関わる複数の神経変性疾患
2013年
- 2013年11/12月号 ダイニン:チームとして強力に作用するモータータンパク質
- 2013年10月号 神経変性:Rhes、SUMO化、ハンチントン病
- 2013年9月号 モノユビキチン化:タンパク質調節のダイナミックなタグ
- 2013年8月号 Ras及びRhoのプレニル化による翻訳後修飾:癌創薬における役割
- 2013年7月号 アクチンが引き起こす膜突起による浸潤:コルタクチン
- 2013年6月号 アクチン修飾と細胞骨格
- 2013年5月号 微小管内部の実体
- 2013年4月号 神経変性におけるTauの多面性
- 2013年3月号 蛍光フィブロネクチンタンパク質を用いた特発性肺線維症の創薬
- 2013年1/2月号 樹状突起棘:発生におけるArf6の役割
2012年
- 2012年11/12月号 ミオシンの小分子モジュレーター
- 2012年10月号 Rhoファミリーパスウェイのユビキチン化と制御
- 2012年9月号 神経変性におけるRac1 GTPaseの機能
- 2012年8月号 上皮間葉転換(EMT)とRhoファミリー低分子量G-タンパク質の関与
- 2012年7月号 チューブリンの多重修飾:グルタミル化とグリシル化
- 2012年6月号 細胞接着のフィブロネクチン制御と原線維形成
- 2012年5月号 アクチン酸化サイクルの機能
- 2012年4月号 トラフィッキング:ArfとCdc42/Racの結合
- 2012年3月号 G-LISAを用いた心臓研究: 糖尿病性心筋症におけるRho経路に関する研究
- 2012年1月/2月号 FtsZ: 新たな抗生剤の標的となるチュ−ブリンホモログ
商品は「研究用試薬」です。人や動物の医療用・臨床診断用・食品用としては使用しないように、十分ご注意ください。
※ 表示価格について














