






















 このページを印刷する
このページを印刷する
記事ID : 13015
Sos/K-Ras 結合を介して Ras シグナル伝達を制御する新しい低分子阻害剤 CYTOSKELETON NEWS 2014年7月号
Sos/K-Ras 結合を介して Ras シグナル伝達を制御する新しい低分子阻害剤
低分子量Gタンパク質 Ras は、不活性型(GDP 結合型)と活性型(GTP 結合型)の間を行き来する GTPase です。Ras は、正常な細胞増殖や分化に関与するシグナル伝達経路において、重要な役割を果たしています1。Ras の3つのアイソフォームである H-Ras、N-Ras、K-Ras は、ヒト腫瘍において発癌作用を示す遺伝子として、30年以上前に同定されました1,2。Ras シグナル伝達の異常は、30% 以上のヒト癌で確認され、肺癌、大腸癌、膵臓癌に多く認められます1,3。特に K-Ras は、癌研究において最も重要な Ras タンパク質であると考えられており、21% 以上のヒト癌に関与しています4。広範囲にわたる研究がなされているにも関わらず、効果的な Ras 阻害剤が同定されていないことから、K-Ras をターゲットとした新薬の開発は非常に困難であると考えられています。現在のところ、グアニンヌクレオチド交換因子である Sos (Son of sevenless)タンパク質の上流をターゲットにした、Ras タンパク質の治療用阻害剤の開発が期待されています。Sos には、2つのヒトアイソフォーム Sos1 と Sos2 がありますが、Sos1 にフォーカスした研究がほとんどです。しかし、すべての論文で Sos1/ Sos2 を介するシグナル伝達が区別されているわけではなく、まとめて「Sos」とされています。非定型 Ras シグナル伝達を制御する薬剤ターゲットの同定について、これまでに得られた知見を紹介します。
Ras を介するシグナル伝達は、一般的に、受容体チロシンキナーゼ(RTK)-Sos-Ras 経路を通じて行われます。増殖因子リガンドが結合すると、膜貫通型 RTK の二量体化及び自己リン酸化が起こります。Sos1 は、増殖因子受容体結合タンパク質2(Grb2)と恒常的に相互作用しています。Sos1/Grb2 複合体は、リン酸化 RTK にリクルートされ、Grb2 の Src homology 2 (SH2) ドメインを介して結合します5-9。RTK/Grb2/Sos1 複合体を形成した Sos は、細胞膜(及び細胞内膜)に位置する不活性型(GDP 結合型)Ras に結合します5。Sos が Ras に結合すると、GDP から GTP への変換が促進され、Ras が活性化されます。活性型である GTP 結合型 Ras は、GTPase 活性化タンパク質(GAPs) を介して、不活性型である GDP 結合型に戻ります(図1)1,6。
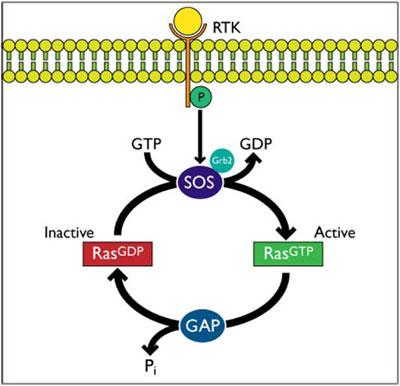
図1 RTK-Grb2-Sos-Ras 経路を介した Ras の活性化
Sos-Ras 相互作用について報告されたのは 20 年前ですが7、最近の研究から、新しい結合ポケット、低分子量化合物、重要な翻訳後修飾が同定され、Sos-Ras 結合の阻害によって作用する、新しい Ras 阻害剤の開発につながることが期待されています。近年発表された4つの論文で、 Sos-Ras 相互作用を変化させたことで生じる興味深い結果が報告されています。
(1)ペプチド HBS3 (水素結合サロゲート[HBS]により合成)は Sos のαへリックスを模倣するように設計されており、Sos-Ras 結合時に Ras のスイッチ領域に挿入されます。in vitro の実験では、Sos-Ras 結合が阻害され、Ras の活性化が有意に低下しました10。
(2)Ras のスイッチ I/II 領域の間に、ユニークなリガンド結合ポケットが同定されました。フラグメントベースドリードディスカバリー(Fragment-Based Lead Discovery [FBLD])により設計された低分子量化合物 DCAI は、このポケットに特異的に結合し、Sos-1 と K-Ras 間の相互作用を防ぐことで K-Ras を阻害します11。
(3)Grb2 のリンカー領域内に位置する Lysine 56 の SUMO化は、Sos1/Grb2 複合体の形成を促進し、おそらく Ras を介すると想定される ERK/MAPK 活性化につながります。反対に、Grb2 の Lysine 56 の SUMO化を阻害すると、Sos1-Grb2 結合が妨げられます12。
(4)K-Ras は、翻訳後修飾として Lysine 104 がアセチル化されると、Sos1を介するヌクレオチド交換が阻害され、負に調節されます。アセチルトランスフェラーゼの活性化又は脱アセチル化酵素の阻害を介して、K-Ras のアセチル化を調節することで、癌の治療につながる可能性があります13。
治療用の K-Ras 阻害剤は、K-Ras 変異体を選択的に阻害することが理想です。変異体と野生型の選択は、変異がエフェクター結合部位から遠位にあるため、困難であることが示されています1。K-Ras の 1アミノ酸残基の変異(12、13、61)は良く知られており、この変異によって K-Ras は恒常的活性化型(GTP 結合)GTPase となります4。 近年、Kevan Shokat 博士の研究グループ(サンフランシスコ カリフォルニア大学 ハワード・ヒューズ医療研究所)が、肺癌患者の 7% に存在する特定の K-Ras 変異体(G12C; グリシン 12 がシステインに置換)で、新しいアロステリックな スイッチ II ポケット(S-IIP)を同定したことで、変異体の選択に関する問題に進展が見られました14。システイン変異に結合し、スイッチ I 及びスイッチ II 領域の機能を阻害する低分子化合物を設計しました。その結果、Sos を介するヌクレオチド交換が妨げられ、K-Ras 変異体の選択的な阻害に成功しています。
Ras の調節機構が明らかになるにつれて、GTPase タンパク質の活性化レベルを高感度で定量的に測定する方法が、ますます重要になっています。サイトスケルトン社では、低分子量Gタンパク質の活性化レベルを広く測定できる、プルダウン法や G-LISA 活性化アッセイなどの、多くの低分子量Gタンパク質ツールを提供しています。プルダウン法は、ビーズに結合したエフェクタータンパク質のドメインを利用して、活性化 GTPase のレベルを測定し、ウエスタンブロットでタンパク質の活性化レベルを測定します。G-LISA 活性化アッセイは、迅速・高感度で、従来のプルダウン法より定量性に優れています。また、Ras 活性化経路の解明に使用できる、低分子量Gタンパク質抗体、活性化剤、阻害剤を提供しています。
参考文献
1. Wang W. et al. 2012. Ras inhibition via direct Ras binding—is there a path forward. Bioorg. Med. Chem. Lett. 22, 5766-5776.
2. Castellano E. and Santos E. 2011. Functional specificity of Ras isoform: so similar but so different. Genes Cancer.2, 216-231.
3. Burns M.C. et al. 2014. Approach for targeting Ras with small molecules that activate SOS-mediated nucleotide exchange. Proc. Natl. Acad. Sci. U.S.A. 111, 3401-3406.
4. Baines A. et al. 2011. Inhibition of Ras for cancer treatment: the search continues. Future Med. Chem. 3, 1787-1808.
5. McKay M.M. and Morrison D.K. 2007. Integrating signals from RTKs to ERK/MAPK. Oncogene. 26, 3113-3121.
6. Innocenti M. et al. 2002. Mechanisms through which sos1 coordinates the activation of Ras and Rac. J. Cell Biol.156, 125-136.
7. Bar-Sagi D. 1994. The Sos (Son of sevenless) protein. Trends Endocrinol. Metab. 5, 165-169.
8. Ceresa B.P. and Pessin J.E. 1998. Insulin regulation of the Ras activation/inactivation cycle. Mol. Cell. Biochem.182, 23-29.
9. Lowenstein E.J. et al. 1992. The SH2 and SH3 domain-containing protein GRB2 links receptor tyrosine kinases to ras signaling. Cell. 70, 431-442.
10. Patgiri A. et al. 2012. An orthosteric inhibitor of the Ras-Sos interaction. Nat. Chem. Biol. 7, 585-587.
11. Maurer T. et al. 2012. Small-molecule ligands bind to a distinct pocket in Ras and inhibit SOS-mediated nucleotide exchange activity. Proc. Natl. Acad. Sci. U.S.A. 109, 5299-5304.
12. Qu Y. et al. 2014. SUMOylation of Grb2 enhances the ERK activity by increasing its binding with Sos1. Mol. Cancer.13, 95
13. Yang M.H. et al. 2012. Regulation of RAS oncogenicity by acetylation. Proc. Natl. Acad. Sci. U.S.A. 109, 10843-10848.
14. Ostrem J. et al. 2013. K-Ras (G12C) inhibitors allosterically control GTP affinity and effector interactions. Nature.503, 548-551.
Ras / Sos 製品
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 | 在庫 |
|---|---|---|---|---|---|
Anti Acetyl Lysine, (Mouse) , 3C6.08.20 |
CYT | AAC01 | 200 UL [2 x 100 μl] |
CYT社 AAC01L 2*100 を参照 |
なし |
| SOS1 Ras GEF protein, Human |
CYT | CS-SOS1-B | 1 MG [1 x 1 mg] |
販売終了 | 販売終了 |
| Raf protein ras binding domain on GST beads, Mouse |
CYT | RF02-A | 1*2 MG |
CYT社 RF02A 2*2 を参照 |
なし |
| Raf protein ras binding domain on GST beads, Mouse |
CYT | RF02-B | 4*2 MG |
CYT社 RF02B 6*2 を参照 |
なし |
| RhoGEF exchange assay |
CYT | BK100 | 1 KIT [60-300 assays] |
¥240,000 | なし |
G-Switch アクチベーター/インヒビター
| 品名 | メーカー | 品番 | 包装 | 希望販売価格 | 在庫 |
|---|---|---|---|---|---|
| Rho Inhibitor I, Clostridium botulinum , Unlabeled |
CYT | CT04-A | 1*20 UG |
¥73,000 | なし |
| Rho Inhibitor I, Clostridium botulinum , Unlabeled |
CYT | CT04-B | 5*20 UG |
¥237,000 | なし |
| Rho Inhibitor I, Clostridium botulinum , Unlabeled |
CYT | CT04-C | 20*20 UG |
お問い合わせ | なし |
| Rho Activator II |
CYT | CN03-A | 3*20 UG |
¥76,000 | なし |
| Rho Activator II |
CYT | CN03-B | 9*20 UG |
¥211,000 | なし |
| Rho/Rac/Cdc42 Activator I |
CYT | CN04-A | 3*20 UG |
¥76,000 | なし |
| Rho/Rac/Cdc42 Activator I |
CYT | CN04-B | 9*20 UG |
¥211,000 | なし |
低分子量Gタンパク質活性化アッセイ/キット
■ CYTOSKELETON NEWS バックナンバー
- 2020年10月号 紡錘体 - 可視化に向けた新規ツール
- 2020年8月号 細胞膜染色用蛍光プローブ
- 2020年4月号 生細胞におけるF-アクチンプローブ
- 2020年3月号 コロナウイルスと細胞骨格
- 2020年2月号 タウ(Tau)の将来性をMapping
- 2020年1月号 Rho GTPaseによる細胞遊走制御
- 2019年12月号 表現型プロファイリング:アクチンに焦点を当てたがん治療
- 2019年11月号 チューブリンの過剰グルタミル化、ミトコンドリア、神経変性
- 2019年9月号 細胞運動性を制御するために相互作用する膜張力とアクチン細胞骨格
- 2019年8月号 Rac1B、がん、およびRac1
- 2019年7月号 Rhoファミリー GTPases、神経可塑性、およびうつ状態
- 2019年6月号 アクチンメチオニン酸化: 動的制御の次の段階
- 2019年5月号 ミクログリアと神経変性疾患
- 2019年2月号 生細胞画像化に対するCNS疾患や障害
2018年
- 2018年12月号 アクチン細胞骨格とメカノトランスダクション(機械的シグナル伝達)
- 2018年11月号 軸索再生と細胞骨格
- 2018年10月号 ニューロンにおける微小管と極性
- 2018年8月号 Rab GTPase と 神経変性
- 2018年7月号 SUMO レスリング: バランスが全て
- 2018年6月号 なぜ K-Ras は発がん特異性を示すのか?
- 2018年5月号 治療標的としてのユビキチンプロテアソームシステム:チューブリンは関与するか?
- 2018年4月号 RhoファミリーGEFと樹状突起スパインの構造的可塑性
- 2018年3月号 βカテニンとTFC/LEF-1の翻訳後修飾による標準的なWntシグナル制御
- 2018年2月号 がん抑制遺伝子p53の翻訳後修飾による機能の調整
- 2018年1月号 自閉スペクトラム症におけるGEF Trioの役割
2017年
- 2017年12月号 プロフィリン: アクチン結合タンパク質の多機能な役割
- 2017年11月号 ミトコンドリアにおけるアセチル化:新たな考え方と治療への応用の可能性
- 2017年9月号 翻訳後修飾のアセチル化による微小管の安定化
- 2017年8月号 神経軸索におけるアクチンリングを基盤とした周期的膜骨格(PMS)
- 2017年7月号 E3ユビキチンリガーゼMdm2によるがん抑制遺伝子p53の翻訳後制御
- 2017年6月号 多能性幹細胞(PSC)での転写因子による翻訳後制御
- 2017年5月号 Arf6 GEF と癌細胞の浸潤・転移
- 2017年4月号 PTEN(Phosphatase and Tensin Homolog)による翻訳後制御
- 2017年3月号 Tau の翻訳後修飾: アルツハイマー病の治療標的
- 2017年2月号 樹状細胞の移動におけるアクチン結合タンパク質とF-アクチン
2016年
- 2016年11月/12月号 GEF を介した GTPase シグナル伝達の低分子阻害剤
- 2016年9月号 FtsZ タンパク質: 抗菌薬の新規ターゲット
- 2016年7月号 翻訳後修飾(PTM)は心臓病において細胞骨格タンパク質を調節する
- 2016年6月号 モータータンパク質キネシンと神経変性
- 2016年5月号 チロシンリン酸化は Rhoファミリー GTPase 活性を調節する
- 2016年4月号 Rac1と糖尿病: ポジティブな役割とネガティブな役割
- 2016年3月号 SUMO化: 細胞骨格タンパク質の機能を調節するレギュレーター
- 2016年1月/2月号 ビメンチン中間径フィラメント: リン酸化による調節
2015年
- 2015年8月号 タンパク質調節に不可欠な翻訳後修飾
- 2015年7月号 アクチン細胞骨格のライブセルイメージング
- 2015年6月号 有糸分裂に関わるタンパク質のSUMO化: 局在と機能
- 2015年5月号 Ras 癌の治療: 5つの有望なターゲット
- 2015年4月号 Ras 依存性の癌で注目される YAP1
- 2015年3月号 増刊号 統合失調症において遺伝子変異により誘導されるアクチン依存のシナプスの変化
- 2015年3月号 Ral GTPase を調節する翻訳後修飾
- 2015年1月/2月号 RhoA のリン酸化はシグナル伝達を調節する
- 2015年1月号 増刊号 微小管を不安定化する suprafenacine: 新規抗癌剤のリード化合物としての可能性
2014年
- 2014年12月号 増刊号 RhoA は心筋細胞におけるアクチン細胞骨格の再構成とグルコース取り込みを仲介する
- 2014年11月号 増刊号 樹状突起の形態形成: ドーパミンD1受容体 および Rho ファミリー GTPase による制御
- 2014年11月/12月号 GTPase 活性化アッセイ: アイソフォームの検出
- 2014年10月号 アルギニンの正電荷を消失させるシトルリン化
- 2014年9月号 キネシンサブドメインの探索
- 2014年9月号 増刊号 アクチン結合タンパク質コフィリンの S-ニトロシル化: 細胞移動に対する影響
- 2014年8月号 増刊号 原発性硬化性胆管炎における N-Ras 発現および活性
- 2014年8月号 SUMO化: 細胞骨格タンパク質を標的とした翻訳後修飾
- 2014年7月号 Sos/K-Ras 結合を介して Ras シグナル伝達を制御する新しい低分子阻害剤
- 2014年6月号 増刊号 頭頸部扁平上皮癌における microRNA-138 による RhoC のダウンレギュレーション
- 2014年6月号 Rho GTPase と活性酸素種: クロストークとフィードバック
- 2014年5月号 ミオシンのアセチル化はサルコメアの構造と機能を調節する
- 2014年4月号 リジンのアセチル化 - 多様な細胞プロセスの制御因子
- 2014年3月号 インテグリンを介したβ-アクチンの酸化還元制御: PDIの出現
- 2014年1/2月号 ダイニン: 一つのモーターが関わる複数の神経変性疾患
2013年
- 2013年11/12月号 ダイニン:チームとして強力に作用するモータータンパク質
- 2013年10月号 神経変性:Rhes、SUMO化、ハンチントン病
- 2013年9月号 モノユビキチン化:タンパク質調節のダイナミックなタグ
- 2013年8月号 Ras及びRhoのプレニル化による翻訳後修飾:癌創薬における役割
- 2013年7月号 アクチンが引き起こす膜突起による浸潤:コルタクチン
- 2013年6月号 アクチン修飾と細胞骨格
- 2013年5月号 微小管内部の実体
- 2013年4月号 神経変性におけるTauの多面性
- 2013年3月号 蛍光フィブロネクチンタンパク質を用いた特発性肺線維症の創薬
- 2013年1/2月号 樹状突起棘:発生におけるArf6の役割
2012年
- 2012年11/12月号 ミオシンの小分子モジュレーター
- 2012年10月号 Rhoファミリーパスウェイのユビキチン化と制御
- 2012年9月号 神経変性におけるRac1 GTPaseの機能
- 2012年8月号 上皮間葉転換(EMT)とRhoファミリー低分子量G-タンパク質の関与
- 2012年7月号 チューブリンの多重修飾:グルタミル化とグリシル化
- 2012年6月号 細胞接着のフィブロネクチン制御と原線維形成
- 2012年5月号 アクチン酸化サイクルの機能
- 2012年4月号 トラフィッキング:ArfとCdc42/Racの結合
- 2012年3月号 G-LISAを用いた心臓研究: 糖尿病性心筋症におけるRho経路に関する研究
- 2012年1月/2月号 FtsZ: 新たな抗生剤の標的となるチュ−ブリンホモログ
商品は「研究用試薬」です。人や動物の医療用・臨床診断用・食品用としては使用しないように、十分ご注意ください。
※ 表示価格について














