






















 このページを印刷する
このページを印刷する
Rhoファミリー GTPases、神経可塑性、およびうつ状態
うつ状態は、個々の認知、情動、動機付け、および生理学的幸福に悪影響を及ぼす様々な症状で構成されています。この不均一な症状および病態群は大うつ病性障害(MDD)と名付けられています。40%もの人々がMDDを患っているものの、薬物療法では十分かつ持続性のある治療的緩和を与えることができません 1,2)。死後のヒト脳の機能的イメージング研究と神経病理学研究より、脳のドーパミン作動性(DAergic)報酬経路にある重要な神経核である側坐核(NAc)と腹側被葢野(VTA)における機能障害が暗示されています 2-6)。VTAはDAニューロンの主な個体群で構成され、NAcを主に神経支配します 2,5)。これらの核における機能障害性神経生理と可塑性は、特に快楽や意欲の喪失(快感消失症)といったMDDのいくつかの症状に寄与すると思われます 2-6)。快感消失症は慢性社会的敗北ストレス(CSDS)を用いて研究され、70%のマウスにおいて抑うつ関連行動(例えば、社会的交流の低減、快感消失の増大、負の体重変化)を誘発(ストレス感受性と呼ばれる)し、残りの30%はこれらの有害な行動変化を起こしません(ストレスに強いと呼ばれる) 2,7)。
VTAのDAergicニューロンとDA受容体1(D1R)- およびD2R-発現NAcニューロンの構造や機能はCSDSの際に再構築されます 2,8-16)。抑うつ様行動を形づくるストレス誘導性快感消失により、D1R発現MSNsへの自発性の興奮性入力が低減するのに対し、D2R発現MSNsは反対の変化を受けます 17,18)。D1R発現(D2Rは発現しない)側坐核ニューロンも樹状突起分枝(形態学的可塑性)の低減を示しますが、樹状突起スパインが興奮性シナプスの原発部位であるため、このことはMSNsにおいて興奮性神経伝達の重要なパラメータとなります 8,10-12,19,20)。
MDDのストレス誘導性モデルは樹状突起棘形態、密度、またはシナプス機能の変化(可塑性など)に関与します 2,12,21,22)。実際、CSDSはNAcにおけるD1R発現MSNsの樹状突起複雑性や樹状突起全長を低減させるものの、D2R発現ではこれが生じません 23)(図1)。樹状突起やスパイン形態におけるこのような変化にはアクチン細胞骨格の同定再構築が必要です。RhoファミリーGTPases(Rho, Rac、およびCdc42など)はスパインなどのアクチン基盤の神経細胞構造の形態形成やリモデリングを制御します 24-28)。一般に、形態形成や発生に対してRhoは阻害を、Rac/Cdc42は促進を行います 2,24,29,30)。上記の知見より、RhoファミリーGTPasesはNAcニューロンにおけるCSDS誘導性ニューロン可塑性にどのような役割を担うのだろうか、といった疑問が自然に湧いてきます。D1R発現MSNsでは、RhoA、そのすぐ下流のエフェクターであるRhoキナーゼ(ROCK)、およびRho GEF Arhgef1の遺伝子発現が著しく増大しています 23)。特に、その他のGTPase遺伝子プロファイルに変化がないことが特徴的です。C3転移酵素とともにRhoAによる側坐核内阻害がなされるとストレス誘導性社会的回避が妨げられ、RhoAの側坐核内活性化により閾値以下のストレス処理が可能となり社会的回避が生じます 23)。同様に、NAcにおけるD1R発現MSCsに対して、低分子の選択的RhoA阻害剤であるRhosinは慢性的なストレス誘導性RhoA活性の増大と興奮性亢進を反転し、自発性の興奮性入力を低減します(図1)。さらに、Rhosinは樹状突起スパイン(興奮性神経伝達の部位)の密度を増強します 31)。RhoAの過活性化を防ぐことで、ニューロン機能や樹状突起構造におけるストレス誘導性欠損への耐性が与えられます 31)(図1)。D1R発現MSNsにおいてストレス誘導性社会的回避と電気生理学的活性変化が同時に生じると、樹状突起分枝の損失、樹状突起全長の減少、および樹状突起に沿って枝分かれ部位数の減少が起こります。RhoAの過活性化とその後のCSDSもまた、全般的な樹状突起形態におけるこれらの有害な変化と関連があります 23)。さらに、コントロールマウスに比べてCSDS処理マウスでは、ROCK阻害剤であるY27632を7日間全身注射した後、同様の社会的行動修復や樹状突起の複雑さが確認されています 23)(図1)。
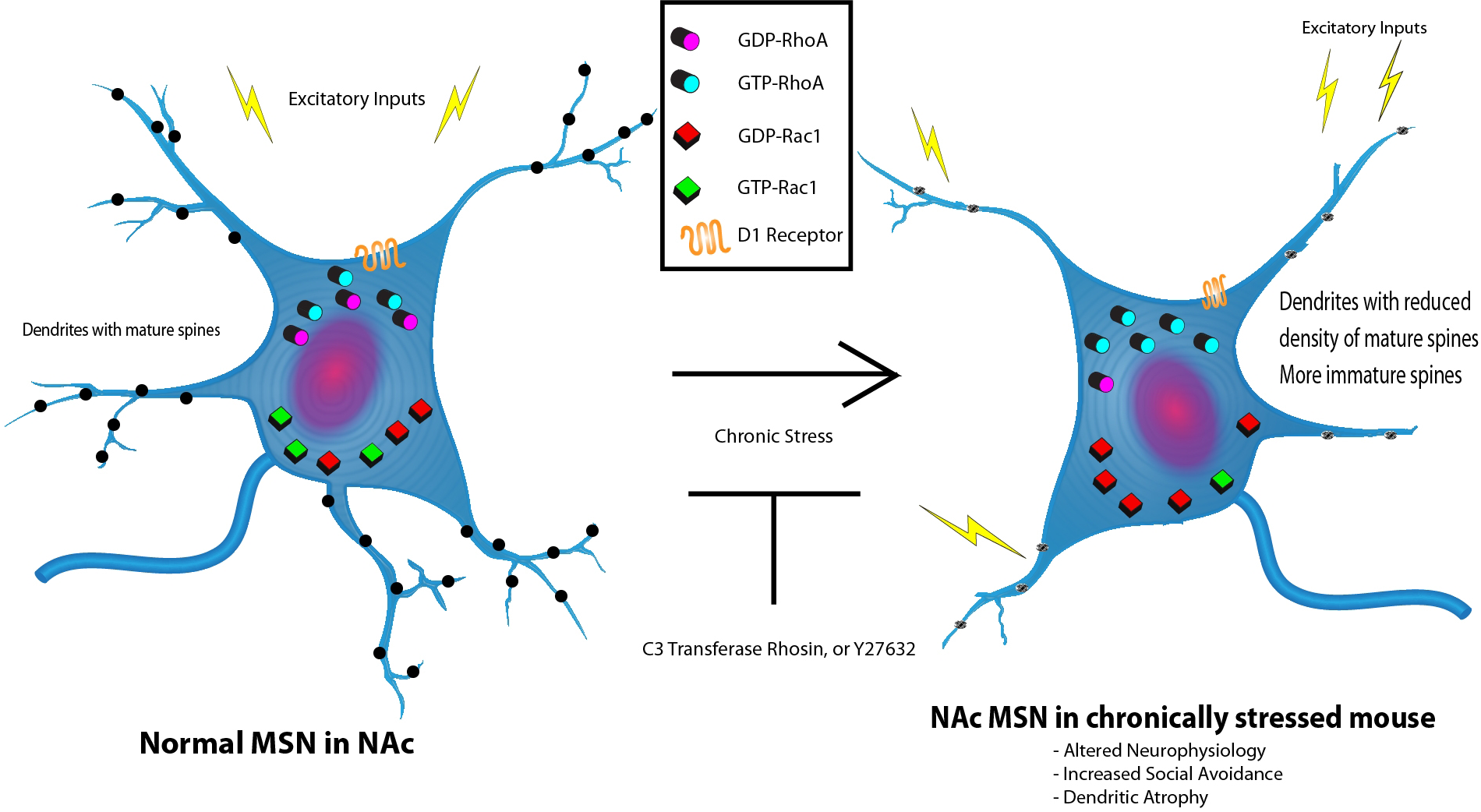
図1 慢性的社会的敗北ストレス(CSDS)に応答した側坐核(NAc)内のドパミン1受容体(D1R)を発現する中型有棘神経細胞(MSNs)における機能障害性可塑性
CSDSに曝露されると、NAcのD1R発現MSNsでは、活性化RhoAのレベル上昇、興奮性入力の低減、興奮性亢進、樹状萎縮(複雑さや成熟スパインの損失、密度低下がみられることもある)、これらのニューロンをもつ動物において社会性回避の増大、および活性化Rac1のレベル低下が見られる。負の神経生理、行動性、および構造的棘変化は、RhoやROCK阻害剤(例えば、C3転移酵素、rhosin、Y27632)により遮断または逆転できた。
NAcにおけるRhoA発現や活性のCSDS介在性増大に反して、他のRhoファミリーGTPaseであるRac1はCSDS後にNAcにおいて特異的に転写および翻訳レベルが下方制御されます 19)。検討したRhoファミリーGTPaseの中でも、Rac1のみがCSDS感受性(回復力のない)マウスにおいてストレス誘導型転写制御を示し、これは社会性回避的行動の増大と相関します 19)(図1)。Rac1転写物レベルも薬物治療していないMDD患者の死後脳のNAcにおいて減少しています 19)。感受性ではあるが回復力のないマウスにおいて、Rac1転写物とタンパク質レベルのCSDS介在性減少はエピジェネティック制御を介しており、Rac1プロモータの許容的アセチル化の減少も示します。さらに、感受性マウスではプロモータのすぐ上流のメチル化が亢進しており、回復力のないマウスではメチル化が減少しています 2,19)。クラス1型HDACsを側坐核内で阻害するとRac1下方制御と付随する社会的回避行動が逆転します。同様に、抑うつした被験者の死後脳由来NAcでは、Rac1遺伝子発現の低減が見られます 19)。Rac1は、NAcのMSNsにおいて、アクチン細胞骨格のコフィリン介在性再構築を介したスパインの形成と成熟(つまり形態形成)に重要です 2,24,28-30)。CSDSに続くRac1 mRNAとタンパク質の下方制御、社会的回避、および快感消失に付随して、側坐核MSNs上に未熟なずんぐりとした樹状突起が形成され、これがコフィリンと共存します 19)。Rac1タンパク質レベルのレスキューと社会的回避行動やこのようなスパイン発生の逆戻りに相関性があることが示されています 19)。
まとめ
MDDを良好に治療するためには、治療耐性、関連する複数の症状や神経生物学的システム、および高率な緩解といった複数の妨害因子を考慮する必要があります 1,2)。定型のものが全ての治療方法に適するとは考え難く、抑うつに関与する全ての神経生物学システムに関連するシグナル伝達カスケードを探究する必要があります。そのため、Cytoskeleton社では、精製済み細胞骨格タンパク質、機能アッセイ、情報伝達試薬、GTPase活性化アッセイ、抗体、生細胞画像化用プローブ、および細胞や組織における翻訳後修飾の内在性レベル定量キットなどをご提供しています。
- Holtzheimer P.E. and Mayberg H.S. 2011. Stuck in a rut: rethinking depression and its treatment. Trends Neurosci. 34, 1-9.
- Fox M.E. and Lobo M.K. 2019. The molecular and cellular mechanisms of depression: a focus on reward circuitry. Mol. Psychiatry. doi: 10.1038/s41380-019-0415-3.
- Drevets W.C. 2001. Neuroimaging and neuropathological studies of depression: implications for the cognitive-emotional features of mood disorders. Curr. Opin. Neurobiol. 11, 240-249.
- Liotti M. and Mayberg H.S. 2001. The role of functional neuroimaging in the neuropsychology of depression. J. Clin. Exp. Neuropsychol. 23, 121-136.
- Nestler E.J. et al. 2002. Neurobiology of depression. Neuron. 34, 13-25.
- Drysdale A.T. et al. 2017. Resting-state connectivity biomarkers define neurophysiological subtypes of depression. Nat. Med. 23, 28-38.
- Czeh B. et al. 2016. Animal models of major depression and their clinical implications. Prog. Neuropsychopharmacol. Biol. Psychiatry. 64, 293-310.
- Muhammad A. et al. 2012. Stress during development alters dendritic morphology in the nucleus accumbens and prefrontal cortex. Neuroscience. 216, 103-109.
- Krishnan V. et al. 2007. Molecular adaptations underlying susceptibility and resistance to social defeat in brain reward regions. Cell. 131, 391-404.
- Christoffel D.J. et al. 2011a. IkappaB kinase regulates social defeat stress-induced synaptic and behavioral plasticity. J. Neurosci. 31, 314-321.
- Christoffel D.J. et al. 2011b. Structural and synaptic plasticity in stress-related disorders. Rev. Neurosci. 22, 535-549.
- Christoffel D.J. et al. 2012. Effects of inhibitor of ΚB kinase activity in the nucleus accumbens on emotional behavior. Neuropsychopharmacology. 37, 2615-2623.
- Cao J.L. Et al. 2010. Mesolimbic dopamine neurons in the brain reward circuit mediate susceptibility to social defeat and antidepressant action. J. Neurosci. 30, 16453-16458.
- Chaudhury D. et al. 2013. Rapid regulation of depression-related behaviours by control of midbrain dopamine neurons. Nature. 493, 532-536.
- Wook K.J. et al. 2016. Essential role of mesolimbic brain-derived neurotrophic factor in chronic social stress-induced depressive behaviors. Biol. Psychiatry. 80, 469-478.
- Qu Y. et al. 2018. Regional differences in dendritic spine density confer resilience to chronic social defeat stress. Acta Neuropsychiatr. 30, 117-122.
- Lim B.K. et al. 2012. Anhedonia requires MC4R-mediated synaptic adaptations in nucleus accumbens. Nature. 487, 183-189.
- Francis T.C. et al. 2015. Nucleus accumbens medium spiny neuron subtypes mediate depression-related outcomes to social defeat stress. Biol. Psychiatry. 77, 212-222.
- Golden S.A. et al. 2013. Epigenetic regulation of RAC1 induces synaptic remodeling in stress disorders and depression. Nat. Med. 19, 337-344.
- Francis T.C. et al. 2017. Molecular basis of dendritic atrophy and activity in stress susceptibility. Mol. Psychiatry. 22, 1512-1519.
- Manji H.K. et al. 2003. Enhancing neuronal plasticity and cellular resilience to develop novel, improved therapeutics for difficult-to-treat depression. Biol. Psychiatry. 53, 707-742.
- Vidal R. et al. 2011. New strategies in the development of antidepressants: towards the modulation of neuroplasticity pathways. Curr. Pharm. Des. 17, 521-533.
- Fox M.E. et al. 2018. Dendritic remodeling of D1 neurons by RhoA/Rho-kinase mediates depression-like behavior. Mol. Psychiatry. DOI: 10.1038/s41380-018-0211-5.
- Nakayama A.Y. et al. 2000. Small GTPases Rac and Rho in the maintenance of dendritic spines and branches in hippocampal pyramidal neurons. J. Neurosci. 20, 5329-5338.
- Luo L. 2002. Actin cytoskeleton regulation in neuronal morphogenesis and structural plasticity. Annu. Rev. Cell Dev. Biol. 18, 601-635.
- Newey S.E. et al. 2005. Rho GTPases, dendritic structure, and mental retardation. J. Neurobiol. 64, 58-74.
- Kiraly D.D. et al. 2010. Synaptic plasticity, a symphony in GEF. ACS Chem. Neurosci. 1, 348-365.
- Tolias K.F. et al. 2011. Control of synapse development and plasticity by Rho GTPase regulatory proteins. Prog. Neurobiol. 94, 133-148.
- Kuhn T.B. et al. 2000. Regulating actin dynamics in neuronal growth cones by ADF/cofilin and rho family GTPases. J. Neurobiol. 44, 126-144.
- Tashiro A. and Yuste R. 2004. Regulation of dendritic spine motility and stability by Rac1 and Rho kinase: evidence for two forms of spine motility. Mol. Cell Neurosci. 26, 429-440.
- Francis T.C. et al. 2019. The selective RhoA inhibitor rhosin promotes stress resiliency through enhancing D1-medium spiny neuron plasticity and reducing hyperexcitability. iol. Psychiatry. 85, 1001-1010















